В нервном центре рефлекса — Студопедия
Трансформацией ритма в нервном центре
152. ПОД ТРАНСФОРМАЦИЕЙ РИТМА ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) направленное распространение возбуждения в ЦНС
2) циркуляцию импульсов в нейронной ловушке
3) беспорядочное распространение возбуждения в ЦНС
Увеличение или уменьшение числа импульсов
153. С УВЕЛИЧЕНИЕМ СИЛЫ РАЗДРАЖИТЕЛЯ ВРЕМЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ
1) не меняется
2) увеличивается
Уменьшается
154. ПРИ УТОМЛЕНИИ ВРЕМЯ РЕФЛЕКСА
1) не меняется
2) уменьшается
Увеличивается
155. В ОСНОВЕ РЕФЛЕКТОРНОГО ПОСЛЕДЕЙСТВИЯ ЛЕЖИТ
1) пространственная суммация импульсов
2) трансформация импульсов
3) последовательная суммация импульсов
Циркуляция импульсов в нейронной ловушке
156. ПОД ДИФФУЗНОЙ ИРРАДИАЦИЕЙ ВОЗБУЖДЕНИЯ ПОНИМАЮТ
2) изменение ритма возбуждения
3) замедленное распространение возбуждения по ЦНС
Ненаправленное распространение возбуждения по ЦНС
157. ПОВЫШАЮЩАЯ ТРАНСФОРМАЦИЯ РИТМА ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ ОБУСЛОВЛЕНА
1) дисперсией возбуждений и низкой лабильностью нервных центров
2) синаптической задержкой
3) утомляемостью нервных центров и дисперсией возбуждений
Дисперсией и мультипликацией возбуждений
158. РОЛЬ СИНАПСОВ ЦНС ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО ОНИ
1) являются местом возникновения возбуждения в ЦНС
2) формируют потенциал покоя нервной клетки
3) проводят токи покоя
Передают возбуждение с нейрона на нейрон
159. В РЕФЛЕКТОРНОЙ ДУГЕ С НАИМЕНЬШЕЙ СКОРОСТЬЮ ВОЗБУЖДЕНИЕ РАСПРОСТРАНЯЕТСЯ ПО ПУТИ
1) афферентному
2) эфферентному
Центральному
160. ЗА ВРЕМЯ РЕФЛЕКСА ПРИНИМАЮТ ВРЕМЯ ОТ НАЧАЛА ДЕЙСТВИЯ РАЗДРАЖИТЕЛЯ ДО
1) конца действия раздражителя
2) достижения полезного приспособительного результата
Появления ответной реакции
161. В ОСНОВЕ ОККЛЮЗИИ ЛЕЖАТ ПРОЦЕССЫ
1) пролонгирования
2) дисперсии
3) мультипликации
Конвергенции
162. ВРЕМЯ РЕФЛЕКСА ЗАВИСИТ ПРЕЖДЕ ВСЕГО
1) от иррадиации возбуждения
2) от физических и химических свойств эффектора
3) от физиологических свойств эффектора
От силы раздражителя и функционального состояния ЦНС
163. ВОЗБУЖДЕНИЕ В НЕРВНОМ ЦЕНТРЕ РАСПРОСТРАНЯЕТСЯ
1) от эфферентного нейрона через промежуточные к афферентному
2) от промежуточных нейронов через эфферентный нейрон к афферентному
3) от промежуточных нейронов через афферентный нейрон к эфферентному
От афферентного нейрона через промежуточные к эфферентному
164. РОЛЬ ЗВЕНА ОБРАТНОЙ АФФЕРЕНТАЦИИ ЗАКЛЮЧАЕТСЯ В ОБЕСПЕЧЕНИИ
1) морфологического соединения нервного центра с эффектором
2) распространения возбуждения от афферентного звена к эфферентному
Оценки результата рефлекторного акта
165. НЕРВНАЯ КЛЕТКА ВЫПОЛНЯЕТ ВСЕ ФУНКЦИИ, КРОМЕ
1) приема информации
2) хранения информации
3) кодирования информации
4) выработки медиатора
Инактивации медиатора
166. ОСНОВНОЙ ФУНКЦИЕЙ ДЕНДРИТОВ ЯВЛЯЕТСЯ
1) проведение возбуждения от тела клетки к эффектору
2) выработка медиатора
Проведение возбуждения к телу нейрона
167. В ЕСТЕСТВЕННЫХ УСЛОВИЯХ ПОТЕНЦИАЛ ДЕЙСТВИЯ В НЕЙРОНЕ ВОЗНИКАЕТ
1) в области дендритов
2) в синапсе
3) в соме нервной клетки
В начальном сегменте аксона
168. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ В ЦНС ОСУЩЕСТВЛЯЕТСЯПРЕИМУЩЕСТВЕННО С УЧАСТИЕМ СИНАПСОВ
1) электрических
2) смешанных
Химических
169. ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ НЕЙРОНА ЗАКЛЮЧАЕТСЯ В
1) посттетанической потенциации
2) связи с другими нейронами посредством отростков
Суммации всех постсинаптических потенциалов, возникающих на мембране нейрона
170. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ ВОЗНИКАЕТ ПРИ ЛОКАЛЬНОЙ
1) гиперполяризации
Деполяризации
171. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ РАЗВИВАЕТСЯ В РЕЗУЛЬТАТЕ ОТКРЫТИЯ НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ КАНАЛОВ ДЛЯ ИОНОВ
1) хлора
2) калия
Натрия
172. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ — ЭТО ЛОКАЛЬНЫЙ ПРОЦЕСС ДЕПОЛЯРИЗАЦИИ, РАЗВИВАЮЩИЙСЯ НА МЕМБРАНЕ
1) аксонного холмика
2) саркоплазматической
3) митохондриальной
4) пресинаптической
Постсинаптической
173. С БОЛЕЕ ВЫСОКОЙ ЧАСТОТОЙ ГЕНЕРИРУЮТ ИМПУЛЬСЫ ТЕ НЕЙРОНЫ, У КОТОРЫХ СЛЕДОВАЯ ГИПЕРПОЛЯРИЗАЦИЯ ДЛИТСЯ
1) 150 мсек
2) 100 мсек
3) 75 мсек
Мсек
174. КОМПЛЕКС СТРУКТУР, НЕОБХОДИМЫХ ДЛЯ ОСУЩЕСТВЛЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ, НАЗЫВАЮТ
1) функциональной системой
2) нервным центром
3) нервно-мышечным препаратом
4) доминантным очагом возбуждения
Рефлекторной дугой
175. ПРИ ДЛИТЕЛЬНОМ РАЗДРАЖЕНИИ КОЖИ ЛАПКИ ЛЯГУШКИ РЕФЛЕКТОРНОЕ ОТДЕРГИВАНИЕ ЛАПКИ ПРЕКРАЩАЕТСЯ ИЗ-ЗА РАЗВИТИЯ УТОМЛЕНИЯ
1) в мышцах лапки
2) в нервно-мышечных синапсах
в нервном центре рефлекса
176. УВЕЛИЧЕНИЕ ЧИСЛА ВОЗБУЖДЕННЫХ НЕЙРОНОВ В ЦНС ПРИ УСИЛЕНИИ РАЗДРАЖЕНИЯ ПРОИСХОДИТ ВСЛЕДСТВИЕ
1) пространственной суммации
2) облегчению
3) окклюзии
Иррадиации
177. РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ ОТ ОДНОГО АФФЕРЕНТНОГО НЕЙРОНА НА МНОГИЕ ИНТЕРНЕЙРОНЫ НАЗЫВАЕТСЯ ПРОЦЕССОМ
1) трансформации ритма
2) пространственной суммации
3) облегчения
4) общего конечного пути
Иррадиации
178. ОДИН МОТОНЕЙРОН МОЖЕТ ПОЛУЧАТЬ ИМПУЛЬСЫ ОТ НЕСКОЛЬКИХ АФФЕРЕНТНЫХ НЕЙРОНОВ В РЕЗУЛЬТАТЕ
1) афферентного синтеза
2) последовательной суммации
3) дивергенции
Конвергенции
179. УСИЛЕНИЕ РЕФЛЕКТОРНОЙ РЕАКЦИИ НЕ МОЖЕТ ВОЗНИКНУТЬ В РЕЗУЛЬТАТЕ
1) торможения рефлекса-антагониста
2) посттетанической потенциации
3) последовательной суммации
4) облегчения
Окклюзии
180. ПОСТТЕТАНИЧЕСКАЯ ПОТЕНЦИАЦИЯ ЗАКЛЮЧАЕТСЯ В УСИЛЕНИИ РЕФЛЕКТОРНОЙ РЕАКЦИИ НА РАЗДРАЖЕНИЕ, КОТОРОМУ ПРЕДШЕСТВОВАЛО
1) торможение нервного центра
2) пространственная суммация импульсов
3) понижающая трансформация импульсов
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА — Студопедия
ВОЗБУЖДЕНИЕ В ЦНС
Выберите один правильный ответ.
151. ЯВЛЕНИЕ ИЗМЕНЕНИЯ КОЛИЧЕСТВА НЕРВНЫХ ИМПУЛЬСОВ В ЭФФЕРЕНТНЫХ ВОЛОКНАХ РЕФЛЕКТОРНОЙ ДУГИ ПО СРАВНЕНИЮ С АФФЕРЕНТНЫМИ ОБУСЛОВЛЕНО
1) рефлекторным последействием
2) наличием доминантного очага возбуждения
3) посттетанической потенциацией
4) трансформацией ритма в нервном центре
152. ПОД ТРАНСФОРМАЦИЕЙ РИТМА ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) направленное распространение возбуждения в ЦНС
2) циркуляцию импульсов в нейронной ловушке
3) беспорядочное распространение возбуждения в ЦНС
4) увеличение или уменьшение числа импульсов
153. С УВЕЛИЧЕНИЕМ СИЛЫ РАЗДРАЖИТЕЛЯ ВРЕМЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ
1) не меняется
2) увеличивается
3) уменьшается
154. ПРИ УТОМЛЕНИИ ВРЕМЯ РЕФЛЕКСА
1) не меняется
2) уменьшается
3) увеличивается
155. В ОСНОВЕ РЕФЛЕКТОРНОГО ПОСЛЕДЕЙСТВИЯ ЛЕЖИТ
1) пространственная суммация импульсов
2) трансформация импульсов
3) последовательная суммация импульсов
4) циркуляция импульсов в нейронной ловушке
156. ПОД ДИФФУЗНОЙ ИРРАДИАЦИЕЙ ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) направленное распространение возбуждения по ЦНС
2) изменение ритма возбуждения
3) замедленное распространение возбуждения по ЦНС
4) ненаправленное распространение возбуждения по ЦНС
157. ПОВЫШАЮЩАЯ ТРАНСФОРМАЦИЯ РИТМА ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ ОБУСЛОВЛЕНА
1) дисперсией возбуждений и низкой лабильностью нервных центров
2) синаптической задержкой
3) утомляемостью нервных центров и дисперсией возбуждений
4)дисперсией и мультипликацией возбуждений
158. РОЛЬ СИНАПСОВ ЦНС ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО ОНИ
1) являются местом возникновения возбуждения в ЦНС
2) формируют потенциал покоя нервной клетки
3) проводят токи покоя
4) передают возбуждение с нейрона на нейрон
159. В РЕФЛЕКТОРНОЙ ДУГЕ С НАИМЕНЬШЕЙ СКОРОСТЬЮ ВОЗБУЖДЕНИЕ РАСПРОСТРАНЯЕТСЯ ПО ПУТИ
1) афферентному
2) эфферентному
3)центральному
160. ЗА ВРЕМЯ РЕФЛЕКСА ПРИНИМАЮТ ВРЕМЯ ОТ НАЧАЛА ДЕЙСТВИЯ РАЗДРАЖИТЕЛЯ ДО
1) конца действия раздражителя
2) достижения полезного приспособительного результата
3) появления ответной реакции
161. В ОСНОВЕ ОККЛЮЗИИ ЛЕЖАТ ПРОЦЕССЫ
1) пролонгирования
2) дисперсии
3) мультипликации
4) конвергенции
162. ВРЕМЯ РЕФЛЕКСА ЗАВИСИТ ПРЕЖДЕ ВСЕГО
1) от иррадиации возбуждения
2) от физических и химических свойств эффектора
3) от физиологических свойств эффектора
4) от силы раздражителя и функционального состояния ЦНС
163. ВОЗБУЖДЕНИЕ В НЕРВНОМ ЦЕНТРЕ РАСПРОСТРАНЯЕТСЯ
1) от эфферентного нейрона через промежуточные к афферентному
2) от промежуточных нейронов через эфферентный нейрон к афферентному
3) от промежуточных нейронов через афферентный нейрон к эфферентному
4) от афферентного нейрона через промежуточные к эфферентному
164. РОЛЬ ЗВЕНА ОБРАТНОЙ АФФЕРЕНТАЦИИ ЗАКЛЮЧАЕТСЯ В ОБЕСПЕЧЕНИИ
1) морфологического соединения нервного центра с эффектором
2) распространения возбуждения от афферентного звена к эфферентному
3) оценки результата рефлекторного акта
165. НЕРВНАЯ КЛЕТКА ВЫПОЛНЯЕТ ВСЕ ФУНКЦИИ, КРОМЕ
1) приема информации
2) хранения информации
3) кодирования информации
4) выработки медиатора
5) инактивации медиатора
166. ОСНОВНОЙ ФУНКЦИЕЙ ДЕНДРИТОВ ЯВЛЯЕТСЯ
1) проведение возбуждения от тела клетки к эффектору
2) выработка медиатора
3)проведение возбуждения к телу нейрона
167. В ЕСТЕСТВЕННЫХ УСЛОВИЯХ ПОТЕНЦИАЛ ДЕЙСТВИЯ В НЕЙРОНЕ ВОЗНИКАЕТ
1) в области дендритов
2) в синапсе
3) в соме нервной клетки
4)в начальном сегменте аксона
168. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ В ЦНС ОСУЩЕСТВЛЯЕТСЯПРЕИМУЩЕСТВЕННО С УЧАСТИЕМ СИНАПСОВ
1) электрических
2) смешанных
3) химических
169. ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ НЕЙРОНА ЗАКЛЮЧАЕТСЯ В
1) посттетанической потенциации
2) связи с другими нейронами посредством отростков
3) суммации всех постсинаптических потенциалов, возникающих на мембране нейрона
170. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ ВОЗНИКАЕТ ПРИ ЛОКАЛЬНОЙ
1) гиперполяризации
2) деполяризации
171. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ РАЗВИВАЕТСЯ В РЕЗУЛЬТАТЕ ОТКРЫТИЯ НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ КАНАЛОВ ДЛЯ ИОНОВ
1) хлора
2) калия
3) натрия
172. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ — ЭТО ЛОКАЛЬНЫЙ ПРОЦЕСС ДЕПОЛЯРИЗАЦИИ, РАЗВИВАЮЩИЙСЯ НА МЕМБРАНЕ
1) аксонного холмика
2) саркоплазматической
3) митохондриальной
4) пресинаптической
5) постсинаптической
173. С БОЛЕЕ ВЫСОКОЙ ЧАСТОТОЙ ГЕНЕРИРУЮТ ИМПУЛЬСЫ ТЕ НЕЙРОНЫ, У КОТОРЫХ СЛЕДОВАЯ ГИПЕРПОЛЯРИЗАЦИЯ ДЛИТСЯ
1) 150 мсек
2) 100 мсек
3) 75 мсек
4) 50 мсек
174. КОМПЛЕКС СТРУКТУР, НЕОБХОДИМЫХ ДЛЯ ОСУЩЕСТВЛЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ, НАЗЫВАЮТ
1) функциональной системой
2) нервным центром
3) нервно-мышечным препаратом
4) доминантным очагом возбуждения
5) рефлекторной дугой
175. ПРИ ДЛИТЕЛЬНОМ РАЗДРАЖЕНИИ КОЖИ ЛАПКИ ЛЯГУШКИ РЕФЛЕКТОРНОЕ ОТДЕРГИВАНИЕ ЛАПКИ ПРЕКРАЩАЕТСЯ ИЗ-ЗА РАЗВИТИЯ УТОМЛЕНИЯ
1) в мышцах лапки
2) в нервно-мышечных синапсах
3) в нервном центре рефлекса
176. УВЕЛИЧЕНИЕ ЧИСЛА ВОЗБУЖДЕННЫХ НЕЙРОНОВ В ЦНС ПРИ УСИЛЕНИИ РАЗДРАЖЕНИЯ ПРОИСХОДИТ ВСЛЕДСТВИЕ
1) пространственной суммации
2) облегчению
3) окклюзии
4) иррадиации
177. РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ ОТ ОДНОГО АФФЕРЕНТНОГО НЕЙРОНА НА МНОГИЕ ИНТЕРНЕЙРОНЫ НАЗЫВАЕТСЯ ПРОЦЕССОМ
1) трансформации ритма
2) пространственной суммации
3) облегчения
4) общего конечного пути
5) иррадиации
178. ОДИН МОТОНЕЙРОН МОЖЕТ ПОЛУЧАТЬ ИМПУЛЬСЫ ОТ НЕСКОЛЬКИХ АФФЕРЕНТНЫХ НЕЙРОНОВ В РЕЗУЛЬТАТЕ
1) афферентного синтеза
2) последовательной суммации
3) дивергенции
4) конвергенции
179. УСИЛЕНИЕ РЕФЛЕКТОРНОЙ РЕАКЦИИ НЕ МОЖЕТ ВОЗНИКНУТЬ В РЕЗУЛЬТАТЕ
1) торможения рефлекса-антагониста
2) посттетанической потенциации
3) последовательной суммации
4) облегчения
5) окклюзии
180. ПОСТТЕТАНИЧЕСКАЯ ПОТЕНЦИАЦИЯ ЗАКЛЮЧАЕТСЯ В УСИЛЕНИИ РЕФЛЕКТОРНОЙ РЕАКЦИИ НА РАЗДРАЖЕНИЕ, КОТОРОМУ ПРЕДШЕСТВОВАЛО
1) торможение нервного центра
2) пространственная суммация импульсов
3) понижающая трансформация импульсов
4) ритмическое раздражение нервного центра
181. ПРОСТРАНСТВЕННАЯ СУММАЦИЯ ИМПУЛЬСОВ ОБЕСПЕЧИВАЕТСЯ
1) дивергенцией возбуждения
2) наличием доминантного очага возбуждения
3) наличием обратной связи
4) конвергенцией возбуждения
182. ДЛЯ НЕЙРОНОВ ДОМИНАНТНОГО ОЧАГА НЕ ХАРАКТЕРНА
1) способность к суммации возбуждений
2) способность к трансформации ритма
3) высокая лабильность
4) инерционность
5) низкая лабильность
183. НЕРВНЫЕ ЦЕНТРЫ НЕ ОБЛАДАЮТ СВОЙСТВОМ
1) пластичности
2) высокой чувствительности к химическим раздражителям
3) способности к суммации возбуждений
4) способности к трансформации ритма
5) двустороннего проведения возбуждений
184. ПЛАСТИЧНОСТЬ СИНАПСОВ ХАРАКТЕРНА
1) только для мотонейронов спинного мозга
2) только для высших отделов ЦНС
3) для любого отдела ЦНС
185. УЧАСТИЕ В РАЗЛИЧНЫХ РЕФЛЕКТОРНЫХ РЕАКЦИЯХ ОДНИХ И ТЕХ ЖЕ ЭФФЕРЕНТНЫХ НЕЙРОНОВ И ЭФФЕКТОРОВ ОБУСЛОВЛЕНО НАЛИЧИЕМ
1) пластичности нервных центров
2) полифункциональности нейронов
3) дивергенции возбуждений
4) проторения пути
5) общего конечного пути
186. ПРЕВЫШЕНИЕ ЭФФЕКТА ОДНОВРЕМЕННОГО ДЕЙСТВИЯ ДВУХ СЛАБЫХ АФФЕРЕНТНЫХ ВОЗБУЖДЕНИЙ
НАД СУММОЙ ИХ РАЗДЕЛЬНЫХ ЭФФЕКТОВ НАЗЫВАЮТ
1) суммацией
2) трансформацией
3) мультипликацией
4) иррадиацией
5) облегчением
187. БОЛЕЕ СЛАБЫЙ ЭФФЕКТ ОДНОВРЕМЕННОГО ДЕЙСТВИЯ ДВУХ СИЛЬНЫХ АФФЕРЕНТНЫХ ВХОДОВ В ЦНС,
ЧЕМ СУММА ИХ РАЗДЕЛЬНЫХ ЭФФЕКТОВ, НАЗЫВАЕТСЯ
1) торможением
2) понижающей трансформацией
3) конвергенцией
4) отрицательной индукцией
5) окклюзией
Установите сответствие.
188.
ПРИНЦИП КООРДИНАЦИОННОЙ ДЕЯТЕЛЬНОСТИ ЦНС…. ЗАКЛЮЧАЕТСЯ
А.2 Облегчение 1. В ослаблении эффекта одновременного
Б.1 Окклюзия действия двух сильных раздражителей
по сравнению с суммой их раздельных эффектов.
2. В превышении эффекта одновременного действия двух слабых раздражителей над суммой их раздельных эффектов.
189.
ПРИНЦИП КООРДИНАЦИОННОЙ ДЕЯТЕЛЬНОСТИ ЦНС…. ЗАКЛЮЧАЕТСЯ
А.1 Общий конечный путь 1. В участии в разных рефлекторных реакциях
Б. 2 Принцип доминанты одних и тех же эфферентных нейронов и эффекторов.
2. В наличии в мозге центра, обладающего повышенной возбудимостью, инертностью и способностью тормозить и суммировать возбуждения других нервных центров.
190.
СВОЙСТВО НЕРВНОГО ЦЕНТРА ….ПРОЯВЛЯЕТСЯ
А.2 Посттетаническая 1. В способности изменять свою
потенциация функцию, расширять функциональные возможности
Б.3 Низкая аккомодационная 2. В усилении рефлекторной реакции после длительного ритмического способность раздражения нервного центра.
3. В способности реагировать на медленно нарастающие по силе раздражители.
191.
КЛЕТКИ ЦНС….ВЫПОЛНЯЮТ ФУНКЦИИ
А.3 Нервные 1. Поглощения избытка медиатора, образования миелиновой оболочки, обеспечения трофики.
Б.1 Глиальные 2. Восприятия энергии раздражителя и трансформации ее в нервный импульс.
3. Приема,обработки, хранения и передачи информации.
192.
ИЗМЕНЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ…. ВОЗНИКАЮТ ВСЛЕДСТВИЕ
А.2 Замедление 1. Посттетанической потенциации.
Б.1 Усиление 2. Утомления нервного центра.
3. Циркуляции импульсов в нейронной ловушке.
193.
ИЗМЕНЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ….ВОЗНИКАЮТ ВСЛЕДСТВИЕ
А.2 Уменьшение времени 1. Циркуляции импульсов в нейронной ловушке.
рефлекса 2. Увеличения силы раздражения.
Б.3 Ослабление ответа 3. Окклюзии.
194.
ИЗМЕНЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ….ВОЗНИКАЮТ ВСЛЕДСТВИЕ
А.3 Усиление рефлекторного 1. Торможения нервного центра.
ответа 2. Циркуляции импульсов в нейронной ловушке
Б.2 Рефлекторное последействие 3. Облегчения.
195.
ИЗМЕНЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ….ВОЗНИКАЮТ ВСЛЕДСТВИЕ
А.1 Замедление 1. Утомления нервного центра.
Б.2 Уменьшение времени 2. Увеличения силы раздражения.
рефлекса 3. Окклюзии.
196.
ЯВЛЕНИЕ, ПРОИСХОДЯЩЕЕ В ЦНС….ОБУСЛОВЛЕНО
А.1 Окклюзия 1. Конвергенцией возбуждений.
Б.4 Понижающая трансфор- 2. Циркуляцией импульсов в нейронной
мация ритма возбуждений ловушке.
В.3 Повышающая трансфор- 3. Дисперсией и иррадиацией возбуждения.
мация ритмов возбуждений 4. Суммацией ВПСП.
197.
ПРИМЕРОМ РЕФЛЕКСА….РЕАКЦИЯ
A.1 Является 1. Сокращение мышц кишки при поступление порции химуса
Б.2 Не является 2. Сокращение мышц кишки после аппликации ацетилхолина.
198.
ПРИМЕРОМ РЕФЛЕКСА….РЕАКЦИЯ
A.1 Является 1. Сужение зрачка при яркой вспышке света.
Б.2 Не является 2. Расширение зрачка при закапывании в глаз атропина (блокатора холинорецепторов).
199.
ЗВЕНО РЕФЛЕКТОРНОЙ ДУГИ….ВЫПОЛНЯЕТ ФУНКЦИИ
А.4 Рецепторное 1. Передает информацию о работе эффектора в кору головного мозга
Б.3 Афферентное 2. Центробежное проведение возбуждения от нервного центра к эффекторной структуре
В.5 Центральное 3. Центростремительное проведение возбуждения от рецепторов к нервному центру.
Г.2 Эфферентное 4. Воспринимает энергию раздражителя и преобразует ее в нервный импульс.
5. Осуществляет анализ и синтез полученной информации.
Определите верны или неверны утверждения и связь между ними.
200. Возбуждение в нейроне раньше всего возникает в области сомы, потому что на мембране тела нейрона происходит суммация только возбуждающих постсинаптических потенциалов.
1) НВН
2) ВВВ
3) ВВН
4) ВНН
5) ННН
201. При повреждении мотонейронов спинного мозга скелетные мышцы конечностей теряют способность сокращаться, потому что мышцы иннервируются дендритами мотонейронов.
1) НВН
2) ННН
3) ВВН
4) ВВВ
5) ВНН
202. При повреждении мотонейронов спинного мозга скелетные мышцы конечностей теряют способность сокращаться , потому что мышцы иннервируются аксонами мотонейронов.
1) НВН
2) ННН
3) ВВН
4) ВНН
5) ВВВ
203. У лягушки с разрушенным спинным мозгом отсутствуют все спинальные рефлексы, потому что рефлекторные акты начинаются с возбуждения нервного центра.
1) НВН
2) ННН
3) ВВН
4) ВВВ
5) ВНН
204. Коленный рефлекс относят к полисинаптическим, потому что в рефлекторной дуге коленного рефлекса
есть один центральный и множество нервно-мышечных синапсов.
1) ВВВ
2) ННН
3) ВВН
4) ВНН
5) НВН
205. Коленный рефлекс относят к моносинаптическим, потому что в структуре коленного рефлекса есть только один центральный синапс.
1) НВН
2) ННН
3) ВВН
4) ВНН
5)ВВВ
206. В рефлекторной дуге возбуждение всегда проводится только в одном направлении , потому что синапсы,
передающие возбуждение от афферентных нейронов к эфферентным, обладают односторонним проведением импульсов.
1) НВН
2) ННН
3) ВВН
4) ВНН
5) ВВВ
207. В рефлекторной дуге возбуждение может проводиться в прямом и обратном направлениях, потому что структура рефлекса имеет звено обратной афферентации.
1) ННН
2) ВНН
3) ВВН
4) ВВВ
5) НВН
208. При поражении передних корешков одного сегмента спинного мозга чувствительность в соответствующем метамере тела полностью не исчезает, а лишь ослабляется, потому что каждый передний корешок спинного мозга иннервирует три метамера тела — свой и два прилежащих к нему.
1) ВВВ
2) ННН
3) ВВН
4) ВНН
5) НВН
209. При поражении передних корешков одного сегмента спинного мозгадвигательная активность в соответствующем метамере тела лишь ослабляется, но не прекращается совсем, потому что каждый метамер иннервируется из трех соседних сегментов спинного мозга.
1) НВН
2) ННН
3) ВВН
4) ВНН
5) ВВВ
210. При поражении одного сегмента спинного мозга двигательная активность в соответствующем метамере тела прекращается, потому что в спинном мозге локализуются мотонейроны скелетных мышц.
1) ВВВ
2) ННН
3) ВВН
4) ВНН
5) НВН
211. При поражении задних корешков одного сегмента спинного мозга чувствительность в соответствующем метамере тела исчезает, потому что задние корешки спинного мозга состоят из афферентных нервных волокон.
1) ВВВ
2) ННН
3) ВВН
4) ВНН
5) НВН
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
⇐ ПредыдущаяСтр 10 из 12Следующая ⇒478. Явление изменения количества нервных импульсов в эфферентных волокнах рефлекторной дуги по сравнению с афферентными обусловлено трансформацией ритма в нервном центре.
479. Под трансформацией ритма возбуждения понимают увеличение или уменьшение числа импульсов.
480. С увеличением силы раздражения время рефлекторной реакции уменьшается.
481. При утомлении время рефлекса увеличивается.
482. В основе рефлекторного последействия лежит циркуляция импульсов в нейронной ловушке.
483. Под диффузной иррадиацией возбуждения понимают беспорядочное распространение возбуждения по ЦНС.
484. Повышающую трансформацию ритма возбуждения в нервной системе обусловливают процессы дисперсия и мультипликация возбуждений.
485. Роль синапсов ЦНС заключается в том, что они передают возбуждение с нейрона на нейрон.
486. В рефлекторной дуге с наименьшей скоростью возбуждение распространяется по центральному пути.
487. За время рефлекса принимают время от начала действия раздражителя до появления ответной реакции.
488. В основе окклюзии лежат процессы конвергенции.
489. Время рефлекса зависит прежде всего от силы раздражителя и функционального состояния ЦНС.
490. Возбуждение в нервном центре распространяется от афферентного нейрона через промежуточные к эфферентному.
491. Роль звена обратной афферентации заключается в обеспечении оценки результата рефлекса.
492. Нервная клетка выполняет все функции, кроме инактивации медиатора.
493. Основной функцией дендритов является передача информации к телу нейрона.
494. Потенциал действия в нейроне возникает в начальном сегменте аксона.
495. Проведение возбуждения в ЦНС осуществляется преимущественно с участием химических синапсов.
496. Интегративная деятельность нейрона заключается в суммацнн всех постсинаптических потенциалов.
497. Возбуждающий постсинаптический потенциал возникает при локальной деполяризации.
498. Возбуждающий постсинаптический потенциал развивается в результате открытия на постсинаптической мембране каналов для ионов натрия.
499. Возбуждающий постсинаптический потенциал — это локальный процесс деполяризации, развивающийся на постсинаптической мембране.
500. С более высокой частотой генерируют импульсы те нейроны, у которых следовая гиперполяризация длится 50 мсек.
501. Комплекс структур, необходимых для осуществления рефлекторной реакции, называют рефлекторной дугой.
502. При длительном раздражении кожи лапки лягушки рефлекторное отдергивание лапки прекращается из-за развития утомления в нервном центре рефлекса.
503. Увеличение числа возбужденных нейронов в ЦНС при усилении раздражения происходит благодаря иррадиации.
504. Возбуждение от одного афферентного нейрона передается на многие мотонейроны благодаря явлению иррадиации.
505. Один мотонейрон может получать импульсы от нескольких афферентных нейронов благодаря конвергенции.
506. Усиление рефлекторной реакции не может возникнуть в результате окклюзии.
507. Посттетаническая потенциация заключается в усилении рефлекторной реакции на раздражение, которому предшествовало ритмическое раздражение нервного центра.
508. Пространственная суммация импульсов обеспечивается конвергенцией возбуждения.
509. Для нейронов доминантного очага не характерна низкая лабильность.
510. Нервные центры не обладают свойством двустороннего проведения возбуждений.
511. Принцип общего конечного пути в координационной деятельности ЦНС действителен для любого ее отдела.
512. Рецепторное звено рефлекторной дуги выполняет функции воспринимает энергию раздражителя и преобразует ее в нервный импульс.
513. Афферентный нерв рефлекторной дуги осуществляет центростремительное проведение возбуждение от рецепторов к нервному центру.
514. Нервный центр осуществляет анализ и синтез полученной информации.
515. Медиатор тормозного нейрона как правило, на постсинаптической мембране не вызывает гиперполяризацию.
516. Время рефлекса в опыте Сеченова увеличивается.
517. В опыте Сеченова разрез мозга проводится между зрительными буграми и вышележащими отделами.
518. При развитии пессимального торможения мембрана нейрона находится в состоянии устойчивой длительной деполяризации.
519. Явление , при котором возбуждение одной мышцы сопровождается торможением центра мышцы-антагониста, называется реципрокным торможением.
520. Торможение было открыто Сеченовым при раздражении зрительных бугров.
521. К специфическим тормозным нейронам относятся клетки Пуркинье н Реншоу.
522. Значение реципрокного торможения заключается в обеспечении координации работы центров-антагонистов.
523. Возникновение ВПСП определяют ионы натрия.
524. Возникновение пессимального торможения вероятно при увеличении частоты импульсов.
525. Пресинаптическое торможение развивается в синапсах аксо-аксональных.
526. Механизм пресинаптического торможения связан с длительной деполяризацией.
527. С точки зрения бинарно-химической теории процесс торможения возникает в результате функционирования специальных синапсов, использующих тормозные медиаторы.
528. Торможение — это процесс, препятствующий возникновению или ослабляющий уже возникшее возбуждение.
529. Явление центрального торможения было открыто Сеченовым И.М.
530. В работе нервных центров торможение необходимо для охраны, регуляции и координации функций.
531. Диффузная иррадиация может быть прекращена в результате торможения.
532. О развитии торможения в опыте Сеченова на лягушке судят по изменению времени спинального рефлекса.
533. Сокращение мышц-сгибателей при одновременном расслаблении мышц- разгибателей возможно в результате реципрокного торможения.
534. Торможение нейронов собственными импульсами, поступающими по коллатералям аксона к тормозным клеткам, называют возвратным.
535. С помощью тормозных вставочных клеток Реншоу осуществляется возвратное торможение.
536. Торможение мотонейронов мышц- антагонистов при сгибании и разгибании конечностей называют реципрокным.
537. При сгибании конечности вставочные тормозные нейроны центра мышц-разгибателей заторможены.
538. Тормозной эффект синапса, расположенного вблизи аксонного холмика, по сравнению с другими участками нейрона более сильный.
539. Развитию торможения нейронов способствует гиперполяризация мембраны аксонного холмика.
540. По своему механизму постсинаптическое торможение может быть и де- и гиперполяризованным.
541. По своему механизму пресинаптическое торможение может быть только деполяризованным.
542. После перерезки ниже продолговатого мозга мышечный тонус значительно уменьшится.
543. Контрактильный тонус при перерезке задних корешков спинного мозга исчезнет.
544. При перерезке между красным мозгом и ядром Дейтерса мышечный тонус разгибателей станет выше тонуса сгибателей.
545. При перерезке передних корешков спинного мозга тонус мышц исчезнет.
546. Влияние красного ядра на ядро Дейтерса является тормозным.
547. Черная субстанция на красное ядро оказывает тормозное влияние.
548. Интрафузальные мышечные волокна иннервируются гамма мотонейронами.
549. Экстрафузальные мышечные волокна иннервируются альфа мотонейронами.
550. Интрафузальные мышечные волокна выполняют функцию обеспечения чувствительности «мышечного веретена» к растяжению.
551. Экстрафузальные мышечные волокна выполняют функцию сокращения мышцы.
552. Тела альфа мотонейронов располагаются в передних рогах спинного мозга.
553. Тела гамма мотонейронов располагаются в передних рогах спинного мозга.
554. Возбуждающие импульсы к ядру Дейтерса поступают преимущественно от рецепторов вестибулярного анализатора.
555. Аппарат Гольджи располагается в сухожилиях мышц.
556. Чувствительные окончания первичных афферентов мышечного веретена находятся в ядерной сумке интрафуэальньк волокон.
557. Быстрое (фазное) движение обеспечивают белые мышечные волокна.
558. Медленное тоническое движение обеспечивают красные мышечные волокна.
559. В рецепции состояния мышцы участвуют мышечные волокна интрафузальные.
560. Возбуждение гамма мотонейронов приведет к сокращению интрафузальных мышечных волокон.
561. Возбуждение рецепторов Гольджи приведет к расслаблению экстрафузальных мышечных волокон.
562. Возбуждение альфа мотонейронов приведет к сокращению экстрафузальных мышечных волокон.
563. Рефлексы, возникающие для поддержании позы при движении, называются статокинетическими.
564. Слабый мышечный тонус наблюдается в эксперименте у спинального животного
565. Повышение мышечного тонуса мышц разгибателей наблюдается у животного бульбарного.
566. При недостаточности мозжечка не наблюдается потеря сознания.
567. Для животных с децеребральной ригидностью не характерно резкое понижение тонуса мышц-разгнбателей.
568. В спинном мозге не замыкаются дуги выпрямительных рефлексов.
569. Симпатический отдел автономной нервной системы осуществляет функции: .активирует деятельность мозга, мобилизует защитные и энергетические ресурсы организма; нервные волокна иннервируют все органы и ткани, в т.ч. и клетки самой нервной системы.
570. Парасимпатический отдел автономной нервной системы осуществляет функции: обеспечивает сохранение гомеостаза возбуждения или торможения регулируемых им органов; нервные волокна не иннервируют скелетные мышцы, матку ,ЦНС и большую часть кровеносных сосудов.
571. Метасимпатический отдел автономной нервной системы осуществляет функции: обеспечивает гомеостаз и управление работой внутренних органов посредством структур, расположенных в нервных узлах самих органов.
572. По механизму передачи возбуждения синапсы бывают химические, электрические и смешанные.
573. Синапсы по функции бывают возбуждающие и тормозные.
574. В основе деятельности ЦНС лежит рефлекторный процесс.
575. Нейроглия выполняет следующие функции: трофическую,, барьерную , фагоцитарную, миэлино- образующую и опорную.
576. По месту замыкания рефлексы бывают мезенцефальные, спинальные, бульбарные, и др.
577. Наличием синапсов в ЦНС обусловлены свойства нервных центров: одностороннее проведение и замедление проведения.
578. Главным образом последействие обусловлено суммацией следовой деполяризации, циркуляцией возбуждения по замкнутым нервным сетям и высокой возбудимостью аксонного холмика.
579. Последействие проявляется в том, что возбуждение продолжается после прекращения раздражения
580. Время сухожильных рефлексов равно 0,01 -0,02 мсек.
581. Явление суммацнн возбуждений в нервных центрах впервые было описано И.М. Сеченовым.
582. Последовательная суммация наблюдается при нанесении на один и тот же рецептор нескольких подпороговых импульсов, следующих друг за другом через короткие интервалы времени.
583. Рецептивное поле — это совокупность рецепторов, раздражение которых вызывает один и тот же рефлекс.
584. Пространственная суммация происходит благодаря возбуждению на мотонейроне нескольких синапсов и суммированию их ВПСП.
585. Морфологическим субстратом иррадиации возбуждения является ветвление многократно дихотомирующих отростков и наличие большого количество вставочных нейронов.
586. Основные принципы распространения возбуждения в ЦНС: иррадиация, мультипликация, дивергенция и конвергенция возбуждения.
587. Иррадиация возбуждения в нервных центрах зависит от силы раздражителя и от функционального состояния нервных центров.
588. Явление облегчения наблюдается когда соседние нейронные, пулы перекрываются периферической каймой.
589. Виды центрального торможения: пресинаптическое, пессимальное, посттетаническое (вслед за возбуждением) и постсинаптическое.
590. Тормозный постсинаптический потенциал обусловлен повышением проницаемости постсинаптической мембраны для ионов К и Cl.
591. Пессимальное торможение обусловлено сильной деполяризацией постсинаптической мембраны и снижением ее проницаемости для ионов Na.
592. Торможение вслед за возбуждением (посттетаническое) обусловлено сильной следовой деполяризацией мембраны.
593. Принципы координационной деятельности НДС: реципрокность, обратная связь, общий конечный путь, доминанта.
594. Цепными рефлексы называют, когда один рефлекторный акт обусловливает возникновение другого.
595. Реципрокность обеспечивается следующими механизмами: последовательная и одновременная индукция, наличием тормозных синапсов, образуемых аксонами нервных клеток на нейронах – антагонистах, постсинаптическим торможением
596. Доминантному очагу присущи следующие черты: повышенная возбудимость, способность к суммированию возбуждения, способность тормозить другие рефлексы, высокая стойкость возбуждения.
597. При участии передних бугров четверохолмия осуществляются зрительные ориентировочные рефлексы.
598. Ядра зрительных бугров функционально делятся на специфические и неспецифические.
599. Таламус выполняет функции: перерабатывает информацию, поступающую от всех рецепторов организма; является центром болевой чувствительности, в котором формируется ощущение боли; принимает участие в формировании ощущений, влечений, эмоциональных состояний.
600. Последовательность передачи возбуждения в рефлекторной дуге: афферентная часть, центральная часть, эфферентная часть.
601. Химические синапсы, в отличие от электрических, характеризуются наличием синаптической задержки, наличием одностороннего проведения и эффективной передачей как возбуждения, так и торможения.
602. Для получения явления децеребрационной ригидности необходимо произвести перерезку между средним и продолговатым мозгом.
603. Симптомы нарушения двигательной функции при удалении мозжечка: атония , астазия, атаксия, астения.
604. Медиаторы, оказывающие тормозное влияние: глицин, гамма — аминомасляная кислота, вещество Р.
605. Псевдо-униполярные нейроны, относятся к афферентным нейронам.
606. В постсинаптической мембране при постсинаптическом торможении происходи гиперполяризация.
607. Пространственная суммация обеспечивается конвергенцией синаптических влияний.
608. Закон Белла-Мажанди доказывается тем, что при перерезке передних корешков на одной стороне происходит полное выключение двигательных реакций, но чувствительность этой стороны сохраняется, а при перерезке задних корешков наблюдается выключение чувствительности.
609. Признаки, характерные для статокинетических рефлексов, в отличие от статических: большая сложность, наличие резких фазных ответов, наличие моносинаптических связей, меньшая скорость реакции.
610. При поражении полосатого тела наблюдаются гиперкинезы.
611. При поражении бледного шара наблюдаются гиперкинезы.
612. Нейросекреторную функцию выполняет гипоталамус.
613. В лимбическую систему мозга входят образования: поясная извилина, гиппокамп, мамиллярные тела, миндалина.
614. Лимбическая система выполняет функции: участвует в формировании мотиваций и эмоций; принимает участие в процессах обучения и памяти.
615. Высший отдел регуляции вегетативной нервной системы локализуется в гипоталамусе.
616. Высший центр регуляции гомеостаза локализуется в гипоталамусе.
617. Низший отдел вегетативной нервной системы локализуется в спинном мозге.
618. ЦНС образуют спинной и головной мозг.
619. Один нейрон, как правило, имеет синаптические связи с тысячами других нейронов.
620. Нервная система обеспечивает связь организма с внешней средой.
621. ЦНС обладает трофической функцией.
622. В основе рефлекторного последействия лежит циркуляция импульсов в замкнутых нейронных цепях.
623. Время рефлекса зависит прежде всего от количества синапсов в рефлекторной дуге.
624. Посттетаническая потенциация заключается в усилении рефлекторной реакции на раздражение, которому предшествовало ритмическое раздражение нервного центра.
625. Принцип общего конечного пути в координационной деятельности ЦНС действителен для любого отдела ЦНС.
626. Пессимальное торможение возникает при увеличении частоты импульсов.
627. Процесс первичного торможения возникает в результате функционирования специальных тормозных нейронов.
628. Союзными называют рефлексы, если один рефлекс усиливает другой.
629. Ритмическими называют рефлексы, при которых последовательно чередуются одни и те же акты.
630. Высший центр координации всех двигательных актов, находящийся под контролем двигательной коры называется мозжечком.
631. Мозжечок принимает участие в регуляции вегетативных (не двигательных) актов.
632. При поражении мозжечка могут отмечаться такие нарушения двигательной сферы, как дизэквилибрия, астения, дистония, атаксия, дизартрия.
633. Дизэквилибрия при поражениях мозжечка – это нарушение равновесия .
634. Астения при поражениях мозжечка – это быстрая утомляемость.
635. Дистония при поражениях мозжечка – это диспропорциональность мышечного тонуса.
636. Дизартрия при поражениях мозжечка – это расстройство речи.
637. Атаксия при поражениях мозжечка – это нарушение величины, скорости и направления движений.
638. Диадохокинез при поражениях мозжечка проявляется невозможностью быстро и точно выполнять симметричные движения.
639. Мозжечок не является органом равновесия.
640. Дефицит функций мозжечка при его поражениях в значительной мере компенсирует кора головного мозга.
641. Зрительные бугры являются высшими подкорковыми чувствительными центрами.
642. Таламус перерабатывает информацию, поступающую от всех рецепторов, является высшим подкорковым центром болевой чувствительности, в котором формируется ощущение боли, принимает участие в формировании ощущений, влечений, эмоциональных состояний.
643. Специфические ядра таламуса, получая чувствительную информацию. от определенного вида рецепторов, адресуют ее в корковые отделы соответствующего анализатора.
644. При возбуждении ядер задней доли гипоталамуса развиваются симпатические функциональные эффекты.
645. Раздражение ядер передней доли гипоталамуса вызывает сужение просвета бронхов, угнетение ЧСС.
646. В гипоталамусе находятся центры всех обмена веществ.
647. Лимбической системе не свойственна функция координация сложных двигательных актов.
648. Главную роль в осуществлении двигательных реакций у высших млекопитающих сразу после рождения играют подкорковые базальные ядра.
649. Главную роль в научении человека ходить играет кора головного мозга.
650. Освоение двигательного рефлекторного акта корой головного мозга человека приводит автоматизации двигательного акта.
651. Центрами, осуществляющими наиболее сложные автоматические движения у человека, являются подкорковые базальные ядра.
652. Двигательные мотонейроны коры головного мозга оказывают тормозное действие на полосатое тело.
653. Полосатое тело тормозит бледный шар.
654. Бледный шар тормозит красное ядро среднего мозга.
655. Кора мозга в зависимости от ее функциональной организации подразделяется на сенсорную, двигательную, ассоциативную.
656. Первичные соматосенсорные зоны коры обеспечивают восприятие простых (элементарных) ощущений.
657. Вторичные соматосенсорные зоны коры обеспечивают формирование ощущений, объединяющих их первичные качества.
658. Вторичные соматосенсорные зоны по отношению к первичным расположены по всей границе первичной зоны.
659. Функциональная асимметрия коры больших полушарий генетически детерминирована.
660. Одностороннее поражение мозга в области задней центральной извилины приведет к одностороннему расстройству всех видов чувствительности.
661. Двустороннее поражение мозга в области задней центральной извилины приведет к полной утрате всех видов чувствительности.
662. В области передней центральной извилины коры мозга находится двигательный корковый центр.
663. В затылочной доле мозга находится зрительный корковый центр.
664. В височной доле коры головного мозга находится слуховой корковый центр.
665. Амплитуда потенциалов, регистрируемых на кожных покровах головы человека при ЭЭГ, колеблется в пределах 5-300 мкВ.
666. Частота потенциалов, регистрируемых с покровов головы человека при ЭЭГ — 0,5-30 Гц.
667. Характеристики альфа ритма ЭЭГ — 8-13 Гц; до 50 мкВ.
668. Характеристики бета ритм ЭЭГ — более 13 Гц;, 20-25 мкВ.
669. Характеристики тета ритм ЭЭГ -4-8 Гц; 100-150 мкВ.
670. Характеристики дельта ритм ЭЭГ — 0,5-3,5 Гц; 200-300 мкВ.
671. Альфа ритм ЭЭГ соответствует состоянию физического и психического покоя.
672. Бета ритм ЭЭГ соответствует умственной работе, эмоциональному напряжению.
673. Тета ритм ЭЭГ соответствует состоянию сна, неглубокого наркоза, гипоксии.
674. Дельта ритм ЭЭГ соответствует состоянию глубокого сна или наркоза.
675. Виды нейронов имеют функциональные особенности: — альфа- мотонейрон — это эфферентный нейрон передних рогов спинного мозга, аксон которых иннервирует экстрафузальные волокна скелетных мышц; — гамма — мотонейрон — это эфферентный нейрон передних рогов спинного мозга, аксон которого иннервирует сократительные элементы интрафузальных волокон; — гигантская клетка Беца — это нейрон моторной зоны коры большого мозга, аксон которого участвует в формировании кортикоспинального или кортикобульбарного трактов; — клетка Реншоу — это тормозный интернейрон спинного мозга, принимающий участие в организации возвратного торможения.
676. Рефлекс Ашнера — Данини. проявляется в урежении сердцебиений при надавливании на глазные яблоки; Геринга- Брейера — в торможении вдоха при растяжении легких; висцеро-висцеральный — в изменении деятельности внутренних органов при. раздражении их интерорецепторов; висцеродермальный — в .изменении деятельности внутренних органов при раздражении определенных участков кожи.
677. Тип нервного волокна и его функциональные особенности: А — это аксоны мотонейронов, иннервирующих скелетные мышцы, и афферентные волокна от мышечных рецепторов, имеющие самую высокую скорость проведения- 120м/сек.; В — это преганглионарные вегетативные волокна со скоростью проведения возбуждения 3-18м/сек; С — это постганглионарные вегетативные волокна и афферентные волокна от некоторых рецепторов тепла, давления, боли, имеющие самую низкую скорость проведения возбуждения (0,5 — 3 м/сек).
678. Согласно принципу Дейла, один нейрон использует во всех своих терминалях только один вид медиатора.
679. По аксону нейрона может распространяться только возбуждение. При суммации ВПСП и ТПСП суммарный итог может быть либо положительным, либо отрицательным.
680. В опыте Сеченова измеряют время спинального рефлекса.
681. Опыт Сеченова проводится на таламической лягушке, потому что для проявления торможения необходимо положить на зрительные бугры кристаллик соли.
682. Торможение спинального рефлекса в опыте Сеченова вызывают раздражением зрительных бугров кристалликом соли.
683. Пресинаптическое торможение очень эффективно при обработке поступающей к нейрону информации, потому что при пресинаптическом торможении возбуждение может быть подавлено избирательно на одном синаптическом входе, не влияя на другие синаптические входы.
684. Рецепторы, чувствительные к серотонину, называют серотонинергическими. Серотонин оказывает и возбуждающее, и тормозное влияние.
685. Для демонстрации роли торможения лягушке вводят стрихнин.
686. Для демонстрации торможения лягушке вводят стрихнин, потому что стрихнин блокирует тормозные синапсы.
687. Для демонстрации роли торможения лягушке вводят стрихнин, потому что после введения стрихнина у лягушки наблюдается диффузная иррадиация возбуждения.
688. Нейрон может находиться в состоянии либо возбуждения, либо торможения.
689. Эфферентный парасимпатический путь имеет двухнейронную структуру. Центры парасимпатического отдела вегетативной нервной системы локализуются в головном мозге.
690. Эфферентный симпатический путь имеет двухнейронную структуру.
691. Преганглионарные симпатические волокна короче постганглионарных. Прегангионарные симпатические нервные волокна относятся к типу В , а постганглинарные — к типу С.
692. Преганглионарные нервные волокна симпатического отдела вегетативной нервной системы относятся к типу В.
693. Интрамуральные эфферентные нейроны сердца — общий конечный путь для парасимпатического и метасимпатического отделов ВНС, потому что они передают возбуждение как от преганглионарных волокон вагуса, так и от интрамуральных вставочных нейронов.
694. Метасимпатическая нервная система осуществляет регуляцию висцеральных органов быстрее , чем симпатическая н парасимпатическая , потому что метасимпатические рефлексы являются местными периферическими.
695. Метасимпатические механизмы регуляции освобождают ЦНС от избыточной информации потому что метасимпатические рефлексы замыкаются вне ЦНС — в интрамуральных ганглиях.
696. Объектом иннервации симпатического отдела вегетативной нервной системы является весь организм. Симпатические нервные волокна образуют сплетения вокруг всех сосудов , приносящих кровь органам и тканям.
697. При одновременном прекращении раздражения симпатических и парасимпатических нервных волокон , идущих к сердцу, эффект симпатического нерва длится дольше, потому что активность холинэстеразы выше активности моноаминоксидазы.
698. В тканях внутренних органов медиатором постганглионарных нервных волокон может быть норадреналин , ацетилхолин , гистамин , потому что действие постганглионарных нервных волокон реализуется через адрено-, холино-, гистаминорецепторы.
699. Норадреналин может вызвать как сужение, так и расширение артериол, потому что эффект норадреналина зависит от типа рецепторов (альфа и бета) , с которыми он взаимодействует.
700. Многие функции внутренних органов (например, двигательная) сохраняются после перерезки симпатических н парасимпатических путей , потому что в стенках этих потому что в стенках этих органов существует метасимпатическая система, включающая нейроны-генераторы.
ВОЗБУЖДЕНИЕ И ТОРМОЖЕНИЕ В ЦНС — Студопедия
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
1. ЯВЛЕНИЕ ИЗМЕНЕНИЯ КОЛИЧЕСТВА НЕРВНЫХ ИМПУЛЬСОВ В ЭФФЕРЕНТНЫХ ВОЛОКНАХ РЕФЛЕКТОРНОЙ ДУГИ ПО СРАВНЕНИЮ С АФФЕРЕНТНЫМИ ОБУСЛОВЛЕНО
1) трансформацией ритма в нервном центре
2) наличием доминантного очага возбуждения
3) посттетанической потенциацией
4) рефлекторным последействием
5) наличием общего конечного пути
2. ПОД ТРАНСФОРМАЦИЕЙ РИТМА ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) направленное распространение возбуждения в ЦНС
2) циркуляцию импульсов в нейронной ловушке
3) увеличение или уменьшение числа импульсов
4) беспорядочное распространение возбуждения в ЦНС
5) увеличение или уменьшение числа синапсов
3. С УВЕЛИЧЕНИЕМ СИЛЫ РАЗДРАЖИТЕЛЯ ВРЕМЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ
1) не меняется
2) увеличивается
3) уменьшается
4) увеличивается до определенного предела
5) либо увеличивается, либо уменьшается
4. ПРИ УТОМЛЕНИИ ВРЕМЯ РЕФЛЕКСА
1) не меняется
2) увеличивается
3) уменьшается
4) уменьшается до определенного предела
5) либо увеличивается, либо уменьшается
5. В ОСНОВЕ РЕФЛЕКТОРНОГО ПОСЛЕДЕЙСТВИЯ ЛЕЖИТ
1) пространственная суммация импульсов
2) циркуляция импульсов в нейронной ловушке
3) последовательная суммация импульсов
4) трансформация импульсов
5) замедленное распространение возбуждения по ЦНС
6. ПОД ДИФФУЗНОЙ ИРРАДИАЦИЕЙ ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) ненаправленное распространение возбуждения по ЦНС
2) изменение ритма возбуждения
3) замедленное распространение возбуждения по ЦНС.
4) направленное распространение возбуждения по ЦНС
5) циркуляцию импульсов в нейронной ловушке
7. ПОВЫШАЮЩАЯ ТРАНСФОРМАЦИЯ РИТМА ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ ОБУСЛОВЛЕНА
1) дисперсией возбуждений и низкой лабильностью нервных центров
2) синаптической задержкой
3) утомляемостью нервных центров и дисперсией возбуждений
4) дисперсией и мультипликацией возбуждений
5) дисперсией и высокой чувствительностью нервных центров
 163. РОЛЬ СИНАПСОВ ЦНС ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО ОНИ
163. РОЛЬ СИНАПСОВ ЦНС ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО ОНИ
1) являются местом возникновения возбуждения в ЦНС
2) формируют потенциал покоя нервной клетки
3) передают возбуждение с нейрона на нейрон
4) проводят токи покоя
5) синтезируют медиаторы
8. В РЕФЛЕКТОРНОЙ ДУГЕ С НАИМЕНЬШЕЙ СКОРОСТЬЮ ВОЗБУЖДЕНИЕ РАСПРОСТРАНЯЕТСЯ ПО ПУТИ
1) афферентному
2) эфферентному
3) центральному
4) обратной афферентации
5) скорость везде одинаковая
9. ЗА ВРЕМЯ РЕФЛЕКСА ПРИНИМАЮТ ВРЕМЯ ОТ НАЧАЛА ДЕЙСТВИЯ РАЗДРАЖИТЕЛЯ ДО
1) конца действия раздражителя
2) появления ответной реакции
3) достижения полезного приспособительного результата
4) возникновения первого потенциала действия
5) все ответы неправильные
10. В ОСНОВЕ ОККЛЮЗИИ ЛЕЖАТ ПРОЦЕССЫ
1) пролонгирования
2) дисперсии
3) мультипликации
4) конвергенции
5) циркуляции импульсов в замкнутых нейронных сетях
11.ВРЕМЯ РЕФЛЕКСА ЗАВИСИТ ПРЕЖДЕ ВСЕГО
1) от силы раздражителя и функционального состояния ЦНС
2) от физических и химических свойств эффектора
3) от физиологических свойств эффектора
4) от иррадиации возбуждения
5) от физических и химических свойств раздражителя
12. ВОЗБУЖДЕНИЕ В НЕРВНОМ ЦЕНТРЕ РАСПРОСТРАНЯЕТСЯ
1) от эфферентных нейронов через промежуточные к афферентным
2) от промежуточных нейронов через эфферентные нейроны к афферентным
3) от афферентных нейронов через промежуточные к эфферентным
4) от промежуточных нейронов через афферентные нейроны к эфферентным
5) все ответы неправильные, т.к. действует закон двустороннего проведения возбуждения
13. РОЛЬ ЗВЕНА ОБРАТНОЙ АФФЕРЕНТАЦИИ ЗАКЛЮЧАЕТСЯ В ОБЕСПЕЧЕНИИ
1) морфологического соединения нервного центра с эффектором
2) распространения возбуждения от афферентного звена к эфферентному
3) оценки результата рефлекторного акта
4) закона двухстороннего распространения возбуждения
5) морфологического соединения эффектора с афферентным нейроном
14. НЕРВНАЯ КЛЕТКА ВЫПОЛНЯЕТ ВСЕ ФУНКЦИИ, КРОМЕ
1) приема информации
2) хранения информации
3) кодирования информации
4) выработки медиатора
5) инактивации медиатора
15. ОСНОВНОЙ ФУНКЦИЕЙ ДЕНДРИТОВ ЯВЛЯЕТСЯ
1) проведение возбуждения к телу нейрона
2) выработка медиатора
3) проведение возбуждения от тела клетки к эффектору
4) инактивация медиатора
5) кодирование и хранение информации
16. В ЕСТЕСТВЕННЫХ УСЛОВИЯХ ПОТЕНЦИАЛ ДЕЙСТВИЯ В НЕЙРОНЕ ВОЗНИКАЕТ
1) в области дендритов
2) в синапсе
3) в соме нервной клетки
4) в начальном сегменте аксона
5) в терминали аксона
16. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ В ЦНС ОСУЩЕСТВЛЯЕТСЯ ПРЕИМУЩЕСТВЕННО С УЧАСТИЕМ СИНАПСОВ
1) электрических
2) химических
3) смешанных
4) всех перечисленных
5) правильного ответа нет
17. ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ НЕЙРОНА ЗАКЛЮЧАЕТСЯ В
1) суммации всех постсинаптических потенциалов, возникающих на мембране нейрона
2) связи с другими нейронами посредством отростков
3) посттетанической потенциации
4) генерации потенциала покоя
5) кодировании и хранении информации
18. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ ВОЗНИКАЕТ ПРИ ЛОКАЛЬНОЙ
1) гиперполяризации
2) деполяризации
3) католической депрессии
4) статической поляризации
5) передаче информации
19. ВОЗБУЖДАЮЩИЙ ПОСТСИИАПТИЧЕСКИЙ ПОТЕНЦИАЛ РАЗВИВАЕТСЯ В РЕЗУЛЬТАТЕ ОТКРЫТИЯ НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ КАНАЛОВ ДЛЯ ИОНОВ
1) хлора
2) натрия
3) калия
4) магния
5) водорода
20. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ — ЭТО ЛОКАЛЬНЫЙ ПРОЦЕСС ДЕПОЛЯРИЗАЦИИ, РАЗВИВАЮЩИЙСЯ НА МЕМБРАНЕ
1) аксонного холмика
2) саркоплазматической
3) митохондриальной
4) пресинаптической
5) постсинаптической
21. С БОЛЕЕ ВЫСОКОЙ ЧАСТОТОЙ ГЕНЕРИРУЮТ ИМПУЛЬСЫ ТЕ НЕЙРОНЫ, У КОТОРЫХ СЛЕДОВАЯ ГИПЕРПОЛЯРИЗАЦИЯ ДЛИТСЯ
1) 50 мсек
2) 100 мсек
3) 75 мсек
4) 150 мсек
5) связи между следовой гиперполяризацией и частотой импульсов нет
22. КОМПЛЕКС СТРУКТУР, НЕОБХОДИМЫХ ДЛЯ ОСУЩЕСТВЛЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ, НАЗЫВАЮТ
1) функциональной системой
2) нервным центром
3) рефлекторной дугой
4) доминантным очагом возбуждения
5) нервно-мышечным препаратом
23. ПРИ ДЛИТЕЛЬНОМ РАЗДРАЖЕНИИ КОЖИ ЛАПКИ ЛЯГУШКИ РЕФЛЕКТОРНОЕ ОТДЕРГИВАНИЕ ЛАПКИ ПРЕКРАЩАЕТСЯ ИЗ-ЗА
РАЗВИТИЯ УТОМЛЕНИЯ
1) в мышцах лапки
2) в нервно-мышечных синапсах
3) в нервном центре рефлекса
4) в звене обратной афферентации
5) в вегетативных ганглиях
24. УВЕЛИЧЕНИЕ ЧИСЛА ВОЗБУЖДЕННЫХ НЕЙРОНОВ В ЦНС ПРИ УСИЛЕНИИ РАЗДРАЖЕНИЯ ПРОИСХОДИТ ВСЛЕДСТВИЕ
1) иррадиации
2) облегчения
3) окклюзии
4) пространственной суммации
5) временной суммации
25. РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ ОТ ОДНОГО АФФЕРЕНТНОГО НЕЙРОНА НА МНОГИЕ ИНТЕРНЕЙРОНЫ НАЗЫВАЕТСЯ ПРОЦЕССОМ
1) трансформации ритма
2) иррадиации
3) облегчения
4) проторения пути
5) пространственной суммации
 26. ОДИН МОТОНЕЙРОН МОЖЕТ ПОЛУЧАТЬ ИМПУЛЬСЫ ОТ НЕСКОЛЬКИХ АФФЕРЕНТНЫХ НЕЙРОНОВ В РЕЗУЛЬТАТЕ
26. ОДИН МОТОНЕЙРОН МОЖЕТ ПОЛУЧАТЬ ИМПУЛЬСЫ ОТ НЕСКОЛЬКИХ АФФЕРЕНТНЫХ НЕЙРОНОВ В РЕЗУЛЬТАТЕ
1) афферентного синтеза
2) последовательной суммации .
3) дивергенции
4) конвергенции
5) трансформации ритма
26. УСИЛЕНИЕ РЕФЛЕКТОРНОЙ РЕАКЦИИ НЕ МОЖЕТ ВОЗНИКНУТЬ В РЕЗУЛЬТАТЕ
1) торможения рефлекса-антагониста
2) посттетанической потенциации
3) последовательной суммации
4) окклюзии
5) облегчения
27. ПОСТТЕТАНИЧЕСКАЯ ПОТЕНЦИАЦИЯ ЗАКЛЮЧАЕТСЯ В УСИЛЕНИИ РЕФЛЕКТОРНОЙ РЕАКЦИИ НА РАЗДРАЖЕНИЕ, КОТОРОМУ ПРЕДШЕСТВОВАЛО
1) ритмическое раздражение нервного центра
2) пространственная суммация импульсов
3) понижающая трансформация ритма импульсов
4) торможение нервного центра
5) все перечисленное
28. ПРОСТРАНСТВЕННАЯ СУММАЦИЯ ИМПУЛЬСОВ ОБЕСПЕЧИВАЕТСЯ
1) дивергенцией возбуждения
2) наличием доминантного очага возбуждения
3) конвергенцией возбуждения
4) наличием обратной связи
5) пространственным облегчением
29. ДЛЯ НЕЙРОНОВ ДОМИНАНТНОГО ОЧАГА ХАРАКТЕРНА
1) способность к суммации возбуждений
2) все перечисленное
3) высокая лабильность
4) инерционность
5) способность к трансформации ритма
30. НЕРВНЫЕ ЦЕНТРЫ ОБЛАДАЮТ
1) пластичностью
2) высокой чувствительностью к химическим раздражителям
3) способностью к суммации возбуждений
4) способностью к трансформации ритма
5) всеми перечисленными свойствами
31. УСИЛЕНИЕ РЕФЛЕКТОРНОЙ РЕАКЦИИ ВОЗНИКАЕТ В РЕЗУЛЬТАТЕ
1) торможения рефлекса-антагониста
2) посттетанической потенциации
3) последовательной суммации
4) облегчения
5) всех перечисленных процессов
32. УЧАСТИЕ В РАЗЛИЧНЫХ РЕФЛЕКТОРНЫХ РЕАКЦИЯХ ОДНИХ И ТЕХ ЖЕ ЭФФЕРЕНТНЫХ НЕЙРОНОВ И ЭФФЕКТОРОВ ОБУСЛОВЛЕНО НАЛИЧИЕМ
1) пластичности нервных центров
2) полифункциональности нейронов
3) общего конечного пути
4) проторения пути
5) дивергенции возбуждений
33. ПРЕВЫШЕНИЕ ЭФФЕКТА ОДНОВРЕМЕННОГО ДЕЙСТВИЯ ДВУХ СЛАБЫХ АФФЕРЕНТНЫХ ВОЗБУЖДЕНИЙ НАД СУММОЙ ИХ РАЗДЕЛЬНЫХ ЭФФЕКТОВ НАЗЫВАЮТ
1) суммацией
2) трансформацией
3) мультипликацией
4) облегчением
5) иррадиацией
34. БОЛЕЕ СЛАБЫЙ ЭФФЕКТ ОДНОВРЕМЕННОГО ДЕЙСТВИЯ ДВУХ СИЛЬНЫХ АФФЕРЕНТНЫХ ВХОДОВ В ЦНС, ЧЕМ СУММА ИХ РАЗДЕЛЬНЫХ ЭФФЕКТОВ, НАЗЫВАЕТСЯ
1) окклюзией
2) понижающей трансформацией
3) конвергенцией
4) отрицательной индукцией
5) торможением
35. ДЛЯ РАЗВИТИЯ ТОРМОЖЕНИЯ В ЦНС НЕОБХОДИМО ВСЕ КРОМЕ
1) нарушения целостности нервного центра
2) энергии АТФ
3) открытия хлорных каналов
4) открытия калиевых каналов
5) медиатора
36. МЕДИАТОР ТОРМОЗНОГО НЕЙРОНА, КАК ПРАВИЛО, НА ПОСТ- СИНАПТИЧЕСКОЙ МЕМБРАНЕ ВЫЗЫВАЕТ
1) статическую поляризацию
2) деполяризацию
3) гиперполяризацию
4) инверсию
5) католическую депрессию
37. ВРЕМЯ РЕФЛЕКСА В ОПЫТЕ И.М.СЕЧЕНОВА
1) не изменяется
2) в этом опыте не определяется
3) увеличивается
4) уменьшается
5) может увеличиваться, может уменьшаться
38. В ОПЫТЕ И.М.СЕЧЕНОВА РАЗРЕЗ МОЗГА ПРОВОДИТСЯ МЕЖДУ
1) грудными и поясничными отделами спинного мозга
2) между зрительными буграми и вышележащими отделами
3) продолговатым и спинным мозгом
4) на любом уровне ЦНС
5) правильного ответа нет
39. ТОРМОЖЕНИЕ БЫЛО ОТКРЫТО И.М. СЕЧЕНОВЫМ ПРИ РАЗДРАЖЕНИИ
1) спинного мозга
2) зрительных бугров
3) коры головного мозга
4) мозжечка
5) продолговатого мозга
40. ПРИ РАЗВИТИИ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ МЕМБРАНА НЕЙРОНА НАХОДИТСЯ В СОСТОЯНИИ
1) статической поляризации
2) гиперполяризации
3) устойчивой длительной деполяризации
4) католической депрессии
5) Реполяризации
41. ЯВЛЕНИЕ, ПРИ КОТОРОМ ВОЗБУЖДЕНИЕ ОДНОЙ МЫШЦЫ СОПРОВОЖДАЕТСЯ ТОРМОЖЕНИЕМ ЦЕНТРА МЫШЦЫ-АНТАГОНИСТА, НАЗЫВАЕТСЯ
1) отрицательной индукцией
2) окклюзией
3) облегчением
4) утомлением
5) реципрокным торможением
42. ТОРМОЖЕНИЕ-ЭТО ПРОЦЕСС
1) локальный
2)распространяющийся, если ТПСП достигает критического уровня
3) всегда распространяющийся
4) может быть локальным, может быть распространяющимся
5) правильного ответа нет
43. К СПЕЦИФИЧЕСКИМ ТОРМОЗНЫМ НЕЙРОНАМ ОТНОСЯТСЯ
1) нейроны черной субстанции и красного ядра среднего
мозга
2) пирамидные клетки коры больших полушарий
3) нейроны ядра Дейтерса продолговатого мозга
4) клетки Пуркинье и Реншоу
5) нейроны солнечного сплетения
44. ЯВЛЕНИЕ СОПРЯЖЕННОГО ТОРМОЖЕНИЯ МОЖНО НАБЛЮДАТЬ
1) в опыте И.М.Сеченова
2) в опыте, когда при развитии одного рефлекса раздражается рецепторное поле антагонистического рефлекса
3) при одновременном раздражении рецепторных полей двух спинальных рефлексов
4) в опыте с односторонним разрушением мозжечка
5) в опыте с введением стрихнина
 45.. ЗНАЧЕНИЕ РЕЦИПРОКНОГО ТОРМОЖЕНИЯ ЗАКЛЮЧАЕТСЯ
45.. ЗНАЧЕНИЕ РЕЦИПРОКНОГО ТОРМОЖЕНИЯ ЗАКЛЮЧАЕТСЯ
1) в обеспечении координации работы центров-антагонистов
2) в освобождении ЦНС от переработки несущественной информации
3) в выполнении защитной функции
4) в проторении пути
5) в формировании общего конечного пути
46. ТПСП ВОЗНИКАЕТ ВСЛЕДСТВИЕ ИЗМЕНЕНИЯ ПРОНИЦАЕМОСТИ МЕМБРАНЫ ДЛЯ ИОНОВ
1) натрия
2) натрия и хлора
3) калия и хлора
4) кальция
5) Н+ и ОН—
47. ВОЗНИКНОВЕНИЕ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ ВЕРОЯТНО
1) при низкой частоте импульсов
2) при секреции тормозных медиаторов
3) при возбуждении вставочных тормозных нейронов
4) при увеличении частоты импульсов
5) при блокаде тормозных каналов
48. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ ОСУЩЕСТВЛЯЕТСЯ ПОСРЕДСТВОМ СИНАПСОВ
1) аксо-соматических
2) аксо-аксональных
3) аксо-дендритных
4) сомато-соматических
5) любых
49. МЕХАНИЗМ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ СВЯЗАН СО СЛЕДУЮЩИМ ЯВЛЕНИЕМ
1) длительной деполяризацией
2) работой К — Na-насоса
3) работой Са-насоса
4) гиперполяризацией
отсутствием раздражителя
50. С ТОЧКИ ЗРЕНИЯ БИНАРНО-ХИМИЧЕСКОЙ ТЕОРИИ ПРОЦЕСС ТОРМОЖЕНИЯ ВОЗНИКАЕТ
1) вследствие инактивации холинэстеразы
2) при функционировании специальных тормозных нейронов, вырабатывающих специальные медиаторы
3) в тех же структурах и с помощью тех же медиаторов,
что и процесс возбуждения
4) при уменьшении синтеза возбуждающего медиатора
5) при блокаде тормозных синапсов
51.. С ТОЧКИ ЗРЕНИЯ УНИТАРНО-ХИМИЧЕСКОЙ ТЕОРИИ ТОРМОЖЕНИЕ ВОЗНИКАЕТ
1) вследствие инактивации холинэстеразы
2) при уменьшении синтеза возбуждающего медиатора
3) в тех же структурах и с помощью тех же медиаторов,
что и процесс возбуждения
4) при функционировании специальных тормозных нейронов, вырабатывающих специальные медиаторы
5) при блокаде тормозных синапсов
52. ЯВЛЕНИЕ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ БЫЛО ОТКРЫТО
1) Ч. Шеррингтоном
2) И.М. Сеченовым
3) И.П. Павловым
4) братьями Вебер
5) Н.Е. Введенским
53. ЯВЛЕНИЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ БЫЛО ОТКРЫТО
1) братьями Вебер
2) Ч.Шеррингтоном
3) И.П.Павловым
4) И.М.Сеченовым
5) Н.Е.Введенским
54. ТОРМОЖЕНИЕ — ЭТО ПРОЦЕСС
1) возникающий в результате утомления нервных клеток
2) приводящий к снижению КУД нервной клетки
3) возникающий в рецепторах при чрезмерно сильных раздражителях
4) препятствующий возникновению возбуждения или ослабляющий уже возникшее возбуждение
5) возникающий в рецепторах при чрезмерно длительном раздражении
55. В РАБОТЕ НЕРВНЫХ ЦЕНТРОВ ТОРМОЖЕНИЕ НЕОБХОДИМО ДЛЯ
1) замыкания дуги рефлексов в ответ на раздражение
2) обеспечения сохранности, регуляции и координации функций
3) объединения клеток ЦНС в нервные центры
4) защиты нейронов от чрезмерного возбуждения
5) всего перечисленного
56.ДИФФУЗНАЯ ИРРАДИАЦИЯ МОЖЕТ БЫТЬ ПРЕКРАЩЕНА В РЕЗУЛЬТАТЕ
1) латерального торможения
2) увеличения силы раздражителя
3) введения стрихнина
4) реципрокного торможения
5) всего перечисленного
57. О РАЗВИТИИ ТОРМОЖЕНИЯ В ОПЫТЕ И.М.СЕЧЕНОВА НА ЛЯГУШКЕ СУДЯТ ПО
1) появлению судорожных сокращений лапок
2) изменению времени спинального рефлекса
3) снижению частоты сердцебиений с последующей остановкой сердца
4) вовлечению в рефлекторный ответ мышц передних
конечностей
5) появлению манежных движений
58. СОКРАЩЕНИЕ МЫШЦ-СГИБАТЕЛЕЙ ПРИ ОДНОВРЕМЕННОМ
РАССЛАБЛЕНИИ МЫШЦ-РАЗГИБАТЕЛЕЙ ВОЗМОЖНО В РЕЗУЛЬТАТЕ
1) активного отдыха
2) облегчения распространения возбуждения
3) реципрокного торможения
4) пессимального торможения
5) отрицательной идукции
59. ТОРМОЖЕНИЕ НЕЙРОНОВ СОБСТВЕННЫМИ ИМПУЛЬСАМИ, ПОСТУПАЮЩИМИ ПО КОЛЛАТЕРАЛЯМ АКСОНА К ТОРМОЗНЫМ КЛЕТКАМ, НАЗЫВАЮТ
1) вторичным
2) реципрокным
3) поступательным
4) латеральным
возвратным
60.. С ПОМОЩЬЮ ТОРМОЗНЫХ ВСТАВОЧНЫХ КЛЕТОК РЕНШОУ
ОСУЩЕСТВЛЯЕТСЯ ТОРМОЖЕНИЕ
1) реципрокное
2) латеральное
3) первичное
4) возвратное
5) пессимальное
61.. ТОРМОЖЕНИЕ МОТОНЕЙРОНОВ МЫШЦ-АНТАГОНИСТОВ
ПРИ СГИБАНИИ И РАЗГИБАНИИ КОНЕЧНОСТЕЙ НАЗЫВАЮТ
1) поступательным
2) реципрокным
3) возвратным
4) латеральным
5) правильного ответа нет
244. ПРИ СГИБАНИИ КОНЕЧНОСТИ ВСТАВОЧНЫЕ ТОРМОЗНЫЕ НЕЙРОНЫ ЦЕНТРА МЫШЦ-РАЗГИБАТЕЛЕЙ ДОЛЖНЫ БЫТЬ
1) возбуждены
2) заторможены
3) в состоянии покоя
4) поочередно в каждом из перечисленных состояний
5) состояние не установлено
62.. ТОРМОЗНОЙ ЭФФЕКТ СИНАПСА, РАСПОЛОЖЕННОГО ВБЛИЗИ АКСОННОГО ХОЛМИКА, ПО СРАВНЕНИЮ С ДРУГИМИ УЧАСТКАМИ НЕЙРОНА, БОЛЕЕ
1) слабый
2) сильный
3) продолжительный
4) сильный и продолжительный
5) слабый, но продолжительный
63.. РАЗВИТИЮ ТОРМОЖЕНИЯ НЕЙРОНОВ СПОСОБСТВУЕТ
1) гиперполяризация мембраны аксонного холмика
2) деполяризация сомы и дендритов
3) деполяризация мембраны аксонного холмика и начального сегмента
4) реполяризация сомы и дендритов
5) реполяризация аксонных холмиков
64.. ПО СВОЕМУ МЕХАНИЗМУ ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ МОЖЕТ БЫТЬ
1) и де-, и гиперполяризационным
2) только гиперполяризационным
3) только деполяризационным
4) только реполяризационным
5) и де-, и реполяризационным
65.. ПО СВОЕМУ МЕХАНИЗМУ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ МОЖЕТ БЫТЬ
1) и де-, и гиперполяризационным
2) только деполяризационным
3) только гиперполяризационным
4) только реполяризационным
5) и де-, и реполяризационным
ВОЗБУЖДЕНИЕ И ТОРМОЖЕНИЕ В ЦНС — Студопедия
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
156. ЯВЛЕНИЕ ИЗМЕНЕНИЯ КОЛИЧЕСТВА НЕРВНЫХ ИМПУЛЬСОВ В ЭФФЕРЕНТНЫХ ВОЛОКНАХ РЕФЛЕКТОРНОЙ ДУГИ ПО СРАВНЕНИЮ С АФФЕРЕНТНЫМИ ОБУСЛОВЛЕНО
1) трансформацией ритма в нервном центре
2) наличием доминантного очага возбуждения
3) посттетанической потенциацией
4) рефлекторным последействием
5) наличием общего конечного пути
157. ПОД ТРАНСФОРМАЦИЕЙ РИТМА ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) направленное распространение возбуждения в ЦНС
2) циркуляцию импульсов в нейронной ловушке
3) увеличение или уменьшение числа импульсов
4) беспорядочное распространение возбуждения в ЦНС
5) увеличение или уменьшение числа синапсов
158. С УВЕЛИЧЕНИЕМ СИЛЫ РАЗДРАЖИТЕЛЯ ВРЕМЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ
1) не меняется
2) увеличивается
3) уменьшается
4) увеличивается до определенного предела
5) либо увеличивается, либо уменьшается
159. ПРИ УТОМЛЕНИИ ВРЕМЯ РЕФЛЕКСА
1) не меняется
2) увеличивается
3) уменьшается
4) уменьшается до определенного предела
5) либо увеличивается, либо уменьшается
160. В ОСНОВЕ РЕФЛЕКТОРНОГО ПОСЛЕДЕЙСТВИЯ ЛЕЖИТ
1) пространственная суммация импульсов
2) циркуляция импульсов в нейронной ловушке
3) последовательная суммация импульсов
4) трансформация импульсов
5) замедленное распространение возбуждения по ЦНС
161. ПОД ДИФФУЗНОЙ ИРРАДИАЦИЕЙ ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) ненаправленное распространение возбуждения по ЦНС
2) изменение ритма возбуждения
3) замедленное распространение возбуждения по ЦНС.
4) направленное распространение возбуждения по ЦНС
5) циркуляцию импульсов в нейронной ловушке
162. ПОВЫШАЮЩАЯ ТРАНСФОРМАЦИЯ РИТМА ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ ОБУСЛОВЛЕНА
1) дисперсией возбуждений и низкой лабильностью нервных центров
2) синаптической задержкой
3) утомляемостью нервных центров и дисперсией возбуждений
4) дисперсией и мультипликацией возбуждений
5) дисперсией и высокой чувствительностью нервных центров
 163. РОЛЬ СИНАПСОВ ЦНС ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО ОНИ
163. РОЛЬ СИНАПСОВ ЦНС ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО ОНИ
1) являются местом возникновения возбуждения в ЦНС
2) формируют потенциал покоя нервной клетки
3) передают возбуждение с нейрона на нейрон
4) проводят токи покоя
5) синтезируют медиаторы
164. В РЕФЛЕКТОРНОЙ ДУГЕ С НАИМЕНЬШЕЙ СКОРОСТЬЮ ВОЗБУЖДЕНИЕ РАСПРОСТРАНЯЕТСЯ ПО ПУТИ
1) афферентному
2) эфферентному
3) центральному
4) обратной афферентации
5) скорость везде одинаковая
165. ЗА ВРЕМЯ РЕФЛЕКСА ПРИНИМАЮТ ВРЕМЯ ОТ НАЧАЛА ДЕЙСТВИЯ РАЗДРАЖИТЕЛЯ ДО
1) конца действия раздражителя
2) появления ответной реакции
3) достижения полезного приспособительного результата
4) возникновения первого потенциала действия
5) все ответы неправильные
166. В ОСНОВЕ ОККЛЮЗИИ ЛЕЖАТ ПРОЦЕССЫ
1) пролонгирования
2) дисперсии
3) мультипликации
4) конвергенции
5) циркуляции импульсов в замкнутых нейронных сетях
167. ВРЕМЯ РЕФЛЕКСА ЗАВИСИТ ПРЕЖДЕ ВСЕГО
1) от силы раздражителя и функционального состояния ЦНС
2) от физических и химических свойств эффектора
3) от физиологических свойств эффектора
4) от иррадиации возбуждения
5) от физических и химических свойств раздражителя
168. ВОЗБУЖДЕНИЕ В НЕРВНОМ ЦЕНТРЕ РАСПРОСТРАНЯЕТСЯ
1) от эфферентных нейронов через промежуточные к афферентным
2) от промежуточных нейронов через эфферентные нейроны к афферентным
3) от афферентных нейронов через промежуточные к эфферентным
4) от промежуточных нейронов через афферентные нейроны к эфферентным
5) все ответы неправильные, т.к. действует закон двустороннего проведения возбуждения
169. РОЛЬ ЗВЕНА ОБРАТНОЙ АФФЕРЕНТАЦИИ ЗАКЛЮЧАЕТСЯ В
ОБЕСПЕЧЕНИИ
1) морфологического соединения нервного центра с эффектором
2) распространения возбуждения от афферентного звена к эфферентному
3) оценки результата рефлекторного акта
4) закона двухстороннего распространения возбуждения
5) морфологического соединения эффектора с афферентным нейроном
170. НЕРВНАЯ КЛЕТКА ВЫПОЛНЯЕТ ВСЕ ФУНКЦИИ, КРОМЕ
1) приема информации
2) хранения информации
3) кодирования информации
4) выработки медиатора
5) инактивации медиатора
171. ОСНОВНОЙ ФУНКЦИЕЙ ДЕНДРИТОВ ЯВЛЯЕТСЯ
1) проведение возбуждения к телу нейрона
2) выработка медиатора
3) проведение возбуждения от тела клетки к эффектору
4) инактивация медиатора
5) кодирование и хранение информации
172. В ЕСТЕСТВЕННЫХ УСЛОВИЯХ ПОТЕНЦИАЛ ДЕЙСТВИЯ В
НЕЙРОНЕ ВОЗНИКАЕТ
1) в области дендритов
2) в синапсе
3) в соме нервной клетки
4) в начальном сегменте аксона
5) в терминали аксона
173. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ В ЦНС ОСУЩЕСТВЛЯЕТСЯ
ПРЕИМУЩЕСТВЕННО С УЧАСТИЕМ СИНАПСОВ
1) электрических
2) химических
3) смешанных
4) всех перечисленных
5) правильного ответа нет
174. ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ НЕЙРОНА ЗАКЛЮЧАЕТСЯ В
1) суммации всех постсинаптических потенциалов, возникающих на мембране нейрона
2) связи с другими нейронами посредством отростков
3) посттетанической потенциации
4) генерации потенциала покоя
5) кодировании и хранении информации
175. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ ВОЗНИКАЕТ ПРИ ЛОКАЛЬНОЙ
1) гиперполяризации
2) деполяризации
3) католической депрессии
4) статической поляризации
5) передаче информации
176. ВОЗБУЖДАЮЩИЙ ПОСТСИИАПТИЧЕСКИЙ ПОТЕНЦИАЛ РАЗВИВАЕТСЯ В РЕЗУЛЬТАТЕ ОТКРЫТИЯ НА ПОСТСИНАПТИЧЕСКОЙ
МЕМБРАНЕ КАНАЛОВ ДЛЯ ИОНОВ
1) хлора
2) натрия
3) калия
4) магния
5) водорода
177. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ —
ЭТО ЛОКАЛЬНЫЙ ПРОЦЕСС ДЕПОЛЯРИЗАЦИИ, РАЗВИВАЮЩИЙСЯ
НА МЕМБРАНЕ
1) аксонного холмика
2) саркоплазматической
3) митохондриальной
4) пресинаптической
5) постсинаптической
178. С БОЛЕЕ ВЫСОКОЙ ЧАСТОТОЙ ГЕНЕРИРУЮТ ИМПУЛЬСЫ ТЕ
НЕЙРОНЫ, У КОТОРЫХ СЛЕДОВАЯ ГИПЕРПОЛЯРИЗАЦИЯ ДЛИТСЯ
1) 50 мсек
2) 100 мсек
3) 75 мсек
4) 150 мсек
5) связи между следовой гиперполяризацией и частотой импульсов нет
179. КОМПЛЕКС СТРУКТУР, НЕОБХОДИМЫХ ДЛЯ ОСУЩЕСТВЛЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ, НАЗЫВАЮТ
1) функциональной системой
2) нервным центром
3) рефлекторной дугой
4) доминантным очагом возбуждения
5) нервно-мышечным препаратом
180. ПРИ ДЛИТЕЛЬНОМ РАЗДРАЖЕНИИ КОЖИ ЛАПКИ ЛЯГУШКИ
РЕФЛЕКТОРНОЕ ОТДЕРГИВАНИЕ ЛАПКИ ПРЕКРАЩАЕТСЯ ИЗ-ЗА
РАЗВИТИЯ УТОМЛЕНИЯ
1) в мышцах лапки
2) в нервно-мышечных синапсах
3) в нервном центре рефлекса
4) в звене обратной афферентации
5) в вегетативных ганглиях
181. УВЕЛИЧЕНИЕ ЧИСЛА ВОЗБУЖДЕННЫХ НЕЙРОНОВ В ЦНС
ПРИ УСИЛЕНИИ РАЗДРАЖЕНИЯ ПРОИСХОДИТ ВСЛЕДСТВИЕ
1) иррадиации
2) облегчения
3) окклюзии
4) пространственной суммации
5) временной суммации
182. РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ ОТ ОДНОГО АФФЕРЕНТНОГО НЕЙРОНА НА МНОГИЕ ИНТЕРНЕЙРОНЫ НАЗЫВАЕТСЯ ПРОЦЕССОМ
1) трансформации ритма
2) иррадиации
3) облегчения
4) проторения пути
5) пространственной суммации
 183. ОДИН МОТОНЕЙРОН МОЖЕТ ПОЛУЧАТЬ ИМПУЛЬСЫ ОТ
183. ОДИН МОТОНЕЙРОН МОЖЕТ ПОЛУЧАТЬ ИМПУЛЬСЫ ОТ
НЕСКОЛЬКИХ АФФЕРЕНТНЫХ НЕЙРОНОВ В РЕЗУЛЬТАТЕ
1) афферентного синтеза
2) последовательной суммации .
3) дивергенции
4) конвергенции
5) трансформации ритма
184. УСИЛЕНИЕ РЕФЛЕКТОРНОЙ РЕАКЦИИ НЕ МОЖЕТ ВОЗНИКНУТЬ В РЕЗУЛЬТАТЕ
1) торможения рефлекса-антагониста
2) посттетанической потенциации
3) последовательной суммации
4) окклюзии
5) облегчения
185. ПОСТТЕТАНИЧЕСКАЯ ПОТЕНЦИАЦИЯ ЗАКЛЮЧАЕТСЯ В УСИЛЕНИИ РЕФЛЕКТОРНОЙ РЕАКЦИИ НА РАЗДРАЖЕНИЕ, КОТОРОМУ
ПРЕДШЕСТВОВАЛО
1) ритмическое раздражение нервного центра
2) пространственная суммация импульсов
3) понижающая трансформация ритма импульсов
4) торможение нервного центра
5) все перечисленное
186. ПРОСТРАНСТВЕННАЯ СУММАЦИЯ ИМПУЛЬСОВ ОБЕСПЕЧИВАЕТСЯ
1) дивергенцией возбуждения
2) наличием доминантного очага возбуждения
3) конвергенцией возбуждения
4) наличием обратной связи
5) пространственным облегчением
187. ДЛЯ НЕЙРОНОВ ДОМИНАНТНОГО ОЧАГА ХАРАКТЕРНА
1) способность к суммации возбуждений
2) все перечисленное
3) высокая лабильность
4) инерционность
5) способность к трансформации ритма
188. НЕРВНЫЕ ЦЕНТРЫ ОБЛАДАЮТ
1) пластичностью
2) высокой чувствительностью к химическим раздражителям
3) способностью к суммации возбуждений
4) способностью к трансформации ритма
5) всеми перечисленными свойствами
189. УСИЛЕНИЕ РЕФЛЕКТОРНОЙ РЕАКЦИИ ВОЗНИКАЕТ В РЕЗУЛЬТАТЕ
1) торможения рефлекса-антагониста
2) посттетанической потенциации
3) последовательной суммации
4) облегчения
5) всех перечисленных процессов
190. УЧАСТИЕ В РАЗЛИЧНЫХ РЕФЛЕКТОРНЫХ РЕАКЦИЯХ ОДНИХ И ТЕХ ЖЕ ЭФФЕРЕНТНЫХ НЕЙРОНОВ И ЭФФЕКТОРОВ ОБУСЛОВЛЕНО НАЛИЧИЕМ
1) пластичности нервных центров
2) полифункциональности нейронов
3) общего конечного пути
4) проторения пути
5) дивергенции возбуждений
191. ПРЕВЫШЕНИЕ ЭФФЕКТА ОДНОВРЕМЕННОГО ДЕЙСТВИЯ
ДВУХ СЛАБЫХ АФФЕРЕНТНЫХ ВОЗБУЖДЕНИЙ НАД СУММОЙ ИХ
РАЗДЕЛЬНЫХ ЭФФЕКТОВ НАЗЫВАЮТ
1) суммацией
2) трансформацией
3) мультипликацией
4) облегчением
5) иррадиацией
192. БОЛЕЕ СЛАБЫЙ ЭФФЕКТ ОДНОВРЕМЕННОГО ДЕЙСТВИЯ
ДВУХ СИЛЬНЫХ АФФЕРЕНТНЫХ ВХОДОВ В ЦНС, ЧЕМ СУММА ИХ
РАЗДЕЛЬНЫХ ЭФФЕКТОВ, НАЗЫВАЕТСЯ
1) окклюзией
2) понижающей трансформацией
3) конвергенцией
4) отрицательной индукцией
5) торможением
217. ДЛЯ РАЗВИТИЯ ТОРМОЖЕНИЯ В ЦНС НЕОБХОДИМО ВСЕ
КРОМЕ
1) нарушения целостности нервного центра
2) энергии АТФ
3) открытия хлорных каналов
4) открытия калиевых каналов
5) медиатора
218. МЕДИАТОР ТОРМОЗНОГО НЕЙРОНА, КАК ПРАВИЛО, НА ПОСТ-
СИНАПТИЧЕСКОЙ МЕМБРАНЕ ВЫЗЫВАЕТ
1) статическую поляризацию
2) деполяризацию
3) гиперполяризацию
4) инверсию
5) католическую депрессию
219. ВРЕМЯ РЕФЛЕКСА В ОПЫТЕ И.М.СЕЧЕНОВА
1) не изменяется
2) в этом опыте не определяется
3) увеличивается
4) уменьшается
5) может увеличиваться, может уменьшаться
220. В ОПЫТЕ И.М.СЕЧЕНОВА РАЗРЕЗ МОЗГА ПРОВОДИТСЯ МЕЖДУ
1) грудными и поясничными отделами спинного мозга
2) между зрительными буграми и вышележащими отделами
3) продолговатым и спинным мозгом
4) на любом уровне ЦНС
5) правильного ответа нет
221. ТОРМОЖЕНИЕ БЫЛО ОТКРЫТО И.М. СЕЧЕНОВЫМ ПРИ РАЗДРАЖЕНИИ
1) спинного мозга
2) зрительных бугров
3) коры головного мозга
4) мозжечка
5) продолговатого мозга
222. ПРИ РАЗВИТИИ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ МЕМБРАНА НЕЙРОНА НАХОДИТСЯ В СОСТОЯНИИ
1) статической поляризации
2) гиперполяризации
3) устойчивой длительной деполяризации
4) католической депрессии
5) Реполяризации
223. ЯВЛЕНИЕ, ПРИ КОТОРОМ ВОЗБУЖДЕНИЕ ОДНОЙ МЫШЦЫ
СОПРОВОЖДАЕТСЯ ТОРМОЖЕНИЕМ ЦЕНТРА МЫШЦЫ-АНТАГОНИСТА, НАЗЫВАЕТСЯ
1) отрицательной индукцией
2) окклюзией
3) облегчением
4) утомлением
5) реципрокным торможением
224. ТОРМОЖЕНИЕ-ЭТО ПРОЦЕСС
1) локальный
2)распространяющийся, если ТПСП достигает критического уровня
3) всегда распространяющийся
4) может быть локальным, может быть распространяющимся
5) правильного ответа нет
225. К СПЕЦИФИЧЕСКИМ ТОРМОЗНЫМ НЕЙРОНАМ ОТНОСЯТСЯ
1) нейроны черной субстанции и красного ядра среднего
мозга
2) пирамидные клетки коры больших полушарий
3) нейроны ядра Дейтерса продолговатого мозга
4) клетки Пуркинье и Реншоу
5) нейроны солнечного сплетения
226. ЯВЛЕНИЕ СОПРЯЖЕННОГО ТОРМОЖЕНИЯ МОЖНО НАБЛЮДАТЬ
1) в опыте И.М.Сеченова
2) в опыте, когда при развитии одного рефлекса раздражается рецепторное поле антагонистического рефлекса
3) при одновременном раздражении рецепторных полей двух спинальных рефлексов
4) в опыте с односторонним разрушением мозжечка
5) в опыте с введением стрихнина
 227. ЗНАЧЕНИЕ РЕЦИПРОКНОГО ТОРМОЖЕНИЯ ЗАКЛЮЧАЕТСЯ
227. ЗНАЧЕНИЕ РЕЦИПРОКНОГО ТОРМОЖЕНИЯ ЗАКЛЮЧАЕТСЯ
1) в обеспечении координации работы центров-антагонистов
2) в освобождении ЦНС от переработки несущественной информации
3) в выполнении защитной функции
4) в проторении пути
5) в формировании общего конечного пути
228. ТПСП ВОЗНИКАЕТ ВСЛЕДСТВИЕ ИЗМЕНЕНИЯ ПРОНИЦАЕМОСТИ МЕМБРАНЫ ДЛЯ ИОНОВ
1) натрия
2) натрия и хлора
3) калия и хлора
4) кальция
5) Н+ и ОН—
229. ВОЗНИКНОВЕНИЕ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ ВЕРОЯТНО
1) при низкой частоте импульсов
2) при секреции тормозных медиаторов
3) при возбуждении вставочных тормозных нейронов
4) при увеличении частоты импульсов
5) при блокаде тормозных каналов
230. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ ОСУЩЕСТВЛЯЕТСЯ
ПОСРЕДСТВОМ СИНАПСОВ
1) аксо-соматических
2) аксо-аксональных
3) аксо-дендритных
4) сомато-соматических
5) любых
231. МЕХАНИЗМ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ СВЯЗАН
СО СЛЕДУЮЩИМ ЯВЛЕНИЕМ
1) длительной деполяризацией
2) работой К — Na-насоса
3) работой Са-насоса
4) гиперполяризацией
5) отсутствием раздражителя
232. С ТОЧКИ ЗРЕНИЯ БИНАРНО-ХИМИЧЕСКОЙ ТЕОРИИ ПРОЦЕСС ТОРМОЖЕНИЯ ВОЗНИКАЕТ
1) вследствие инактивации холинэстеразы
2) при функционировании специальных тормозных нейронов, вырабатывающих специальные медиаторы
3) в тех же структурах и с помощью тех же медиаторов,
что и процесс возбуждения
4) при уменьшении синтеза возбуждающего медиатора
5) при блокаде тормозных синапсов
233. С ТОЧКИ ЗРЕНИЯ УНИТАРНО-ХИМИЧЕСКОЙ ТЕОРИИ ТОРМОЖЕНИЕ ВОЗНИКАЕТ
1) вследствие инактивации холинэстеразы
2) при уменьшении синтеза возбуждающего медиатора
3) в тех же структурах и с помощью тех же медиаторов,
что и процесс возбуждения
4) при функционировании специальных тормозных нейронов, вырабатывающих специальные медиаторы
5) при блокаде тормозных синапсов
234. ЯВЛЕНИЕ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ БЫЛО ОТКРЫТО
1) Ч. Шеррингтоном
2) И.М. Сеченовым
3) И.П. Павловым
4) братьями Вебер
5) Н.Е. Введенским
235. ЯВЛЕНИЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ БЫЛО ОТКРЫТО
1) братьями Вебер
2) Ч.Шеррингтоном
3) И.П.Павловым
4) И.М.Сеченовым
5) Н.Е.Введенским
236. ТОРМОЖЕНИЕ — ЭТО ПРОЦЕСС
1) возникающий в результате утомления нервных клеток
2) приводящий к снижению КУД нервной клетки
3) возникающий в рецепторах при чрезмерно сильных раздражителях
4) препятствующий возникновению возбуждения или ослабляющий уже возникшее возбуждение
5) возникающий в рецепторах при чрезмерно длительном раздражении
237. В РАБОТЕ НЕРВНЫХ ЦЕНТРОВ ТОРМОЖЕНИЕ НЕОБХОДИМО ДЛЯ
1) замыкания дуги рефлексов в ответ на раздражение
2) обеспечения сохранности, регуляции и координации функций
3) объединения клеток ЦНС в нервные центры
4) защиты нейронов от чрезмерного возбуждения
5) всего перечисленного
238.ДИФФУЗНАЯ ИРРАДИАЦИЯ МОЖЕТ БЫТЬ ПРЕКРАЩЕНА В РЕЗУЛЬТАТЕ
1) латерального торможения
2) увеличения силы раздражителя
3) введения стрихнина
4) реципрокного торможения
5) всего перечисленного
239. О РАЗВИТИИ ТОРМОЖЕНИЯ В ОПЫТЕ И.М.СЕЧЕНОВА НА ЛЯГУШКЕ СУДЯТ ПО
1) появлению судорожных сокращений лапок
2) изменению времени спинального рефлекса
3) снижению частоты сердцебиений с последующей остановкой сердца
4) вовлечению в рефлекторный ответ мышц передних
конечностей
5) появлению манежных движений
240. СОКРАЩЕНИЕ МЫШЦ-СГИБАТЕЛЕЙ ПРИ ОДНОВРЕМЕННОМ
РАССЛАБЛЕНИИ МЫШЦ-РАЗГИБАТЕЛЕЙ ВОЗМОЖНО В РЕЗУЛЬТАТЕ
1) активного отдыха
2) облегчения распространения возбуждения
3) реципрокного торможения
4) пессимального торможения
5) отрицательной идукции
241. ТОРМОЖЕНИЕ НЕЙРОНОВ СОБСТВЕННЫМИ ИМПУЛЬСАМИ,
ПОСТУПАЮЩИМИ ПО КОЛЛАТЕРАЛЯМ АКСОНА К ТОРМОЗНЫМ
КЛЕТКАМ, НАЗЫВАЮТ
1) вторичным
2) реципрокным
3) поступательным
4) латеральным
5) возвратным
242. С ПОМОЩЬЮ ТОРМОЗНЫХ ВСТАВОЧНЫХ КЛЕТОК РЕНШОУ
ОСУЩЕСТВЛЯЕТСЯ ТОРМОЖЕНИЕ
1) реципрокное
2) латеральное
3) первичное
4) возвратное
5) пессимальное
243. ТОРМОЖЕНИЕ МОТОНЕЙРОНОВ МЫШЦ-АНТАГОНИСТОВ
ПРИ СГИБАНИИ И РАЗГИБАНИИ КОНЕЧНОСТЕЙ НАЗЫВАЮТ
1) поступательным
2) реципрокным
3) возвратным
4) латеральным
5) правильного ответа нет
244. ПРИ СГИБАНИИ КОНЕЧНОСТИ ВСТАВОЧНЫЕ ТОРМОЗНЫЕ
НЕЙРОНЫ ЦЕНТРА МЫШЦ-РАЗГИБАТЕЛЕЙ ДОЛЖНЫ БЫТЬ
1) возбуждены
2) заторможены
3) в состоянии покоя
4) поочередно в каждом из перечисленных состояний
5) состояние не установлено
245. ТОРМОЗНОЙ ЭФФЕКТ СИНАПСА, РАСПОЛОЖЕННОГО ВБЛИЗИ АКСОННОГО ХОЛМИКА, ПО СРАВНЕНИЮ С ДРУГИМИ УЧАСТКАМИ НЕЙРОНА, БОЛЕЕ
1) слабый
2) сильный
3) продолжительный
4) сильный и продолжительный
5) слабый, но продолжительный
246. РАЗВИТИЮ ТОРМОЖЕНИЯ НЕЙРОНОВ СПОСОБСТВУЕТ
1) гиперполяризация мембраны аксонного холмика
2) деполяризация сомы и дендритов
3) деполяризация мембраны аксонного холмика и начального сегмента
4) реполяризация сомы и дендритов
5) реполяризация аксонных холмиков
247. ПО СВОЕМУ МЕХАНИЗМУ ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ МОЖЕТ БЫТЬ
1) и де-, и гиперполяризационным
2) только гиперполяризационным
3) только деполяризационным
4) только реполяризационным
5) и де-, и реполяризационным
248. ПО СВОЕМУ МЕХАНИЗМУ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ МОЖЕТ БЫТЬ
1) и де-, и гиперполяризационным
2) только деполяризационным
3) только гиперполяризационным
4) только реполяризационным
5) и де-, и реполяризационным
Вопрос 9. Рецептивное поле рефлекса, время рефлекса, его зависимость от силы раздражения.
Рефлекс — ответная реакция организма на раздражение из внешней или внутренней среды, осуществляющаяся при участии ЦНС. Вся нервная деятельность складывается из рефлексов различной степени сложности, т. е. является отраженной, вызванной внешним поводом, внешним толчком. Путь, по которому проходит нервный импульс от рецептора до эффектора (действующий орган), называется рефлекторной дугой.
В рефлекторной дуге различают пять звеньев: 1) рецептор; 2) чувствительное волокно, про-водящее возбуждение к центрам; 3) нервный центр, где происходит переключение возбуждения с чувствительных клеток на двигательные; 4) двигательное волокно, передающее нервные импульсы на периферию; 5) действующий орган — мышца или железа. Для осуществления любого рефлекса необходима целостность всех звеньев рефлекторной дуги. Нарушение хотя бы одного из них ведет к исчезновению рефлекса.
Любое раздражение, воспринимаемое рецептором, кодируется в нервный импульс и в таком виде по чувствительным волокнам направляется в ЦНС. Здесь эта информация перерабатывается, отбирается и передается на двигательные нервные клетки, которые посылают нервные импульсы к рабочим органам — мышцам, железам и вызывают тот или иной приспособительный акт — движение или секрецию.
Во время ответной реакции возбуждаются рецепторы рабочего органа и от них в ЦНС поступают импульсы — информация о достигнутом результате. Живой организм, как любая саморегулирующаяся система, работает по принципу обратной связи. Афферентные импульсы, осуществляющие обратную связь, либо усиливают и уточняют реакцию, если она не достигла цели, либо прекращают ее. Таким образом, рефлекс осуществляется не рефлекторной дугой, а рефлекторным кольцом; рефлекс заканчивается по достижении результата.
Время рефлекса. Время, прошедшее от момента нанесения раздражения до ответа на него, называется временем рефлекса (латентный период). Оно слагается из времени, необходимого для возбуждения рецепторов, проведения возбуждения по чувствительным волокнам, ЦНС, двигательным волокнам, и, наконец, скрытого (латентного) периода возбуждения рабочего органа. Большая часть времени уходит на проведение возбуждения через нервные центры — центральное время рефлекса. Это объясняется тем, что в синапсах ЦНС происходит замедление проведения возбуждения, так называемая синаптическая задержка. Чем меньше нейронов входит в состав рефлекторной дуги, тем короче время рефлекса. Поэтому сухожильные рефлексы, возникающие при растяжении сухожилия, имеющие двухнейронную дугу, наиболее быстрые. Их время составляет всего 19-23 мс, тогда как время рефлекса моргания, возникающего при раздражении глаза, равно 50-200 мс. Наибольшим является время вегетативных рефлексов.
Время рефлекса зависит от силы раздражения и возбудимости ЦНС.
Рецептивное поле рефлекса. Каждый рефлекс можно вызвать только с определенного рецептивного поля. Анатомическая область, при раздражении которой вызывается данный рефлекс, носит название рецептивного поля рефлекса. Например, рефлекс сосания возникает при раздражении губ ребенка, рефлекс сужения зрачка — при освещении сетчатки, коленный рефлекс (разгибание голени) — при легком ударе по сухожилию ниже надколенника.
Нервный центр. Каждый рефлекс имеет свою локализацию в ЦНС, т. е. тот ее участок, который необходим для его осуществления. При разрушении соответствующего участка рефлекс отсутствует. Однако выяснилось, что для регуляции рефлекса, его точности недостаточно первичного, или главного, центра, а необходимо участие и высших отделов ЦНС, включая кору большого мозга.
Только при целостности ЦНС сохраняется совершенство нервной деятельности. Нервным центром называется совокупность нервных клеток, расположенных в различных отделах ЦНС, необходимая для осуществления рефлекса и достаточная для его регуляции. Так, если у животного удалить кору полушарий большого мозга, то дыхание сохраняется, так как первичный дыхательный центр находится в продолговатом мозге. Однако во время работы не будет точного соответствия вентиляции легких потребностям организма в кислороде, так как для тонкой регуляции деятельности дыхательного центра необходим не только ствол мозга, но и кора больших полушарий.
Особенности нервных центров. Характерными особенностями нервных центров, отличающими их от нервных волокон, являются быстрая утомляемость, очень высокий обмен веществ, т. е. высокая потребность в кислороде и питательных веществах, и избирательная чувствительность к некоторым ядам. Вследствие этих особенностей нарушения кровоснабжения и изменения температуры тела прежде всего сказываются на функции ЦНС.
Возбуждением называют нервный процесс, который либо вызывает деятельность органа, либо усиливает существующую.
Под торможением понимают такой нервный процесс, который ослабляет либо прекращает деятельность или препятствует ее возникновению. Взаимодействие этих двух активных процессов лежит в основе нервной деятельности.
При сокращении группы мышц тормозятся центры мышц-антагонистов. Так, при сгибании руки или ноги центры мышц-разгибателей затормаживаются. Если бы этого не произошло, то возникла бы механическая борьба мышц, судороги, а не приспособительные двигательные акты.
При раздражении чувствительного нерва, вызывающего сгибательный рефлекс, импульсы направляются к центрам мышц-сгибателей и через тормозные клетки Рёншоу — к центрам мышц-разгибателей. В первых вызывают процесс возбуждения, а во вторых — торможения. В ответ возникает координированный, согласованный рефлекторный акт — сгибательный рефлекс.
Понятие о доминанте. В ЦНС под влиянием тех или иных причин может возникнуть очаг повышенной возбудимости, который обладает свойством притягивать к себе возбуждения с других рефлекторных дуг и тем самым усиливать свою активность и тормозить другие нервные центры. Это явление носит название доминанты. Доминанта относится к числу основных закономерностей в деятельности ЦНС. Она может возникнуть под влиянием различных причин: голода, жажды, инстинкта самосохранения или размножения.
Важным фактором координации рефлексов является наличие в ЦНС функциональной субординации, т. е. определенного соподчинения между ее отделами, возникшего в процессе длительной эволюции. Нервные центры и рецепторы головы «авангардной» части тела, прокладывающей путь организму в окружающей среде, развиваются быстрее. Высшие отделы ЦНС приобретают способность изменять активность и направление
деятельности нижележащих отделов. Нет функций в организме, которые бы не поддавались решающему регулирующему влиянию коры больших полушарий.
Классификация рефлексов.
По биологическому значению рефлексы подразделяются на пищевые, оборонительные, ориентировочные (ознакомление с изменяющимися условиями среды), половые (продолжение рода).
По роду рецепторов, с которых они возникают, рефлексы делятся на экстероцептивные, возникающие с рецепторов, воспринимающих раздражения из внешней среды: световые, звуковые, вкусовые, тактильные и др.; интероцептивные, возникающие с рецепторов внутренних органов: механо-, термо-, осмо- и хеморецепторов сосудов и внутренних органов, и проприоцептивные — с рецепторов, находящихся в мышцах, сухожилиях, связках.
В зависимости от рабочего органа, участвующего в ответной реакции, рефлексы подразделяются на двигательные, секреторные, сосудистые.
По местонахождению главного нервного центра, необходимого для осуществления рефлекса, они делятся на спинальные, например мочеиспускание, дефекация; бульбарные (продолговатый мозг): кашель, чиханье, рвота; мезэнцефальные (средний мозг): выпрямление тела, ходьба; диэнцефальные (промежуточный мозг) — терморегуляторные; корковые — условные рефлексы.
В зависимости от продолжительности различают фазные и тонические рефлексы. Тонические рефлексы длительные, продолжаются часами, например рефлекс стояния. Любое животное может стоять часами благодаря длительному сокращению мышц. Все позные рефлексы относятся к тоническим. Они фиксируют определенное положение тела, а на их фоне разыгрываются другие, короткие, фазные рефлексы, обеспечивающие все виды рабочих, спортивных и других движений.
По сложности рефлексы можно разделить на простые и сложные. Расширение зрачка в ответ на затемнение глаза, разгибание ноги в ответ на легкий удар по сухожилию — это простые рефлексы. Примерами сложных рефлексов служат регуляция сердечно-сосудистой системы, процесс пищеварения. В этих случаях конец одного рефлекса служит раздражителем для возникновения другого. Возникают так называемые цепные рефлексы, протекание которых очень демонстративно можно проследить на примере процесса пищеварения. Произвольное проталкивание комка пищи к задней стенке глотки вызывает раздражение ее рецепторов — возникает рефлекс глотания. Пища попадает в пищевод и вызывает его сокращение, продвигающее пищевой комок ко входу в желудок. Раздражение нижней части пищевода приводит к открытию кардинального жома желудка и поступлению пищи в желудок, а последнее вызывает отделение желудочного сока и т. д. Весь процесс пищеварения — сложная цепь рефлексов.
По принципу эффекторной иннервации рефлексы можно разделить на скелетно-моторные, или соматические (обеспечивающие двигательные акты скелетной мускулатуры), и вегетативные (функции внутренних органов).
В зависимости от того, являются ли рефлексы врожденными или приобретенными в процессе индивидуальной жизни, И. П. Павлов подразделял их на безусловные (врожденные) и условные (приобретенные).
РЕФЛЕКС КАК ЭЛЕМЕНТАРНЫЙ МЕХАНИЗМ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ — Студопедия.Нет
5-1. Рефлекс – это ответная реакция организма на:
1) изменение внешней среды
2) *изменение внешней и внутренней среды, осуществляемая с участием центральной
нервной системы в ответ на раздражение рецепторов
3) раздражение нервного центра спинного или головного мозга
4) изменение внутренней среды
5) раздражение афферентных или эфферентных проводящих путей
5-2. Рефлекторная дуга – это:
1) структурно-функциональная единица ЦНС, состоящая из рецепторов и
исполнительного органа
2) путь, связывающий между собой центральную нервную систему и исполнительный
орган
3) путь, связывающий рецепторы с нервным центром
4) *путь нервных импульсов от рецептора к исполнительному органу
5) путь, связывающий между собой нейроны ЦНС
5-3. Рецепторное звено рефлекторной дуги выполняет функцию:
1) центробежное проведение возбуждения от нервного центра к исполнительной
структуре
2) центростремительное проведение возбуждения от рецепторов к нервному центру,
частотно-спектральное перекодирование
3) *воспринимает действие раздражителя, преобразует его энергию в рецепторный
потенциал и кодирует свойства раздражителей
4) осуществляет анализ и синтез полученной информации, перекодирование информации
и выработку команды
5) осуществляет координацию деятельности эффектора
5-4. Афферентное звено рефлекторной дуги выполняет функции:
1) центробежное проведение возбуждения от нервного центра к исполнительной
структуре
2) *центростремительное проведение возбуждения от рецепторов к нервному центру,
частотно-спектральное перекодирование
3) воспринимает действие раздражителя, преобразует его энергию в рецепторный
потенциал и кодирует свойства раздражителей
4) осуществляет анализ и синтез полученной информации, перекодирование информации
и выработку команды
5) осуществляет координацию деятельности эффектора
5-5. Центральное звено рефлекторной дуги выполняет функции:
1) центробежное проведение возбуждения от нервного центра к исполнительной структуре
2) центростремительное проведение возбуждения от рецепторов к нервному центру, частотно-
спектральное перекодирование
3) воспринимает энергию раздражителя, преобразует ее в рецепторный потенциал и кодирует
свойства раздражителей
4) *осуществляет анализ и синтез полученной информации, перекодирование информации и
выработку команды
5) воспринимает рецепторный потенциал и преобразует его в потенциал действия
5-6. Обратная афферентация – это:
1) центробежное проведение возбуждения от нервного центра к исполнительной структуре
2) центростремительное проведение возбуждения от рецепторов к нервному центру
3) *информация о результате рефлекса, поступающая от рецепторов исполнительного органа
4) анализ и синтез афферентной импульсации
5) восприятие энергии раздражителя
5-7. Если полностью выключить одно из звеньев рефлекторной дуги, то рефлекс:
1) осуществляется
2) *не осуществляется
3) осуществляется только при сверхпороговом раздражении
4) осуществляется нерегулярно
5) осуществляется при наличии обратных связей
5-8. Причиной одностороннего проведения возбуждения в рефлекторной дуге являются особенности:
1) проведения возбуждения по афферентным волокнам
2) проведения возбуждения по эфферентным волокнам
3) *проведения возбуждения в синапсах
4) проведения возбуждения в соме нейрона
5) взаимодействия между нервными центрами
5-9. За латентное (скрытое) время рефлекса принимают время от начала действия раздражителя до:
1) конца действия раздражителя
2) возбуждения нервного центра
3) *появления ответной реакции исполнительного органа
4) появления обратной афферентации
5) завершения рефлекторной реакции
5-10. В рефлекторной дуге обычно наибольшее время задержки проводимого возбуждения имеется в:
1) афферентном звене
2) эфферентном звене
3) *центральном звене
4) звене обратной афферентации
5) задержки проведения возбуждения не наблюдается
5-11. Для собственных рефлексов характерно, что:
1) *рецепторы и эффектор находятся в пределах одной физиологической системы
2) рецепторы и эффектор находятся в разных физиологических системах
3) отсутствует звено обратной афферентации
4) нерегулярная обратная афферентация
5) не наблюдается центральной задержки проведения возбуждения
5-12. Главная часть (ядро) нервного центра в отличие от вспомогательных частей центра:
1) увеличивает адаптивные возможности регулируемой функции
2) *её поражение полностью выключает регулируемую функцию
3) имеет преимущественно полимодальные нейроны
4) тормозит деятельность окружающих нейронов
5) имеет недостаточное количество синаптических контактов от афферентов
5-13. Пластичность нервных центров – это способность:
1) *изменять свое функциональное назначение
2) суммировать приходящее возбуждение и тормозить рядом лежащие центры
3) трансформировать ритм возбуждения
4) к облегчению
5) к окклюзии
5-14. Наибольшей пластичностью обладают:
1) спинальные центры
2) стволовые центры
3) *корковые центры
4) базальные ядра
5) проводящие пути
5-15. Повышающую трансформацию ритма возбуждения в нервном центре обуславливает:
1) низкая лабильность эфферентных нейронов
2) синаптическая задержка
3) утомляемость центра
4) *мультипликации возбуждений
5) конвергенция возбуждений
5-16. Утомляемость нервных центров по сравнению с нервными волокнами:
1) *более высокая
2) более низкая
3) одинаковая
4) не меняется в зависимости от функционального состояния
5) нет правильного ответа
5-17. Функциональное значение реверберации (циркуляции) возбуждения в нервных центрах:
1) *продление времени возбуждения и формирование памяти
2) ослабление возбуждения
3) создание реципрокных отношений в центре
4) торможение возбуждения
5) мультипликация возбуждений
5-18. Избирательно ограничивает отдельные входы поступлении информации к нейрону:
1) *пресинаптическое торможение
2) постсинаптическое торможение
3) возбуждающий постсинаптический потенциал
4) реципрокное торможение
5) сеченовское торможение
5-19. Возвратное торможение:
1) *предупреждает перевозбуждение мотонейронов
2) создает тонус покоя мотонейронов
3) создает реципрокные отношения между мотонейронами
4) вызывает длительную деполяризацию афферентов
5) обеспечивает иррадиацию возбуждения в ЦНС
5-20. Для реципрокного торможения характерно:
1) возбуждение центра тормозит этот же центр через клетки Реншоу
2) *возбуждение нейронов одного центра тормозит возбуждение центра антагонистического
рефлекса
3) возбужденный центр окружает себя зоной торможения
4) латеральное вытормаживание распространения возбуждения
5) длительная деполяризация афферентов
5-21. Латеральное (окружающее) торможение выполняет функцию:
1) подавляет возбуждение вызвавшего его центра
2) *концентрирует возбуждение в данном центре и ограничивает его иррадиацию
3) вызывает иррадиацию возбуждения от данного центра к другим
4) создает реципрокные отношения
5) обеспечивает реверберацию импульсов в нейронной сети типа «нейронной ловушки»
Как быстро ваш мозг реагирует на раздражители
Коленная рефлекторная дуга — это спинномозговой рефлекс, и схема изображена выше. На этом изображении показано, как сенсорный (афферентный) нейрон передает информацию через ганглий дорсального корешка в спинной мозг; где сигнал разделяется на два разных пути. Первый — двигательный нейрон (эфферент), ведущий обратно к четырехглавой мышце. Когда мотонейрон четверной мышцы получает информацию, он срабатывает и заставляет вашу голень подпрыгивать в воздухе.Второй сигнал от сенсорного нейрона проходит к интернейрону, который посылает сигнал на двигательный нейрон (эфферент), ведущий к подколенному сухожилию. Этот сигнал говорит вашему подколенному сухожилию расслабиться, чтобы на четырехглавую мышцу не действовала отрицательная сила при ее сокращении. Оба сигнала работают вместе, и все это происходит в спинном мозге, не доходя до головного мозга. Ему никогда не нужен мозг.
Вы можете спросить, чем отличается дуга рефлекса колена и футболист, сражающийся с приближающимся мячом.Оба ли не рефлексы? Хотя может показаться, что футболист, ведущий переговоры о приближающемся мяче, является простым быстрым рефлексом, на самом деле это симфония сотен тысяч нейронов, работающих вместе для выработки сознательного решения. Ловит ли игрок мяч, уворачивается или отбивает мяч? Этот выбор вызывает реакцию.
Когда футболист понимает, что мяч летит к нему, появляется визуальная информация, которую необходимо обработать, и принять решение относительно правильного образа действий.Затем мозгу необходимо посылать множество сигналов различным мышцам. Ноги начинают двигаться, руки могут перемещаться перед лицом, а глаза могут закрываться, как и многие другие процессы. Это работа многих нейронов, а также многочисленных систем и цепей в мозгу, и, более того, вы можете тренировать и улучшать свои навыки с помощью практики. Так со временем вы станете лучше в спорте.
Как и вся наука, история открытия времени реакции своеобразна. Голландский физиолог Ф.К. Дондерс в 1865 году начал думать о времени реакции человека и о том, можно ли его измерить. До его исследований ученые считали, что психические процессы человека слишком быстры, чтобы их можно было измерить. Это предположение было доказано с помощью Чарльза Уитстона, английского ученого и изобретателя. В 1840 году Уитстон изобрел устройство, очень похожее на его раннее изобретение телеграфной системы, которое регистрировало скорость артиллерийских снарядов. Дондерс использовал это устройство, чтобы измерить время, прошедшее с момента удара по стопе пациента до момента, когда пациент нажал кнопку.На кнопку приходилось нажимать левой или правой рукой, соответствующей левой или правой ноге, получившей удар. В его исследовании проверялось 2 условия: в первом пациент заранее знал, какую ногу следует нанести электрошоку; в другом состоянии пациент не знал. Дондерс обнаружил задержку в 1/15 секунды между пациентами, которые знали, какую ногу следует наносить током, и пациентами, которые не знали. Примечательно, что это был первый отчет об измерении человеческого разума!
Эти усилия продолжаются и сегодня, когда совершенствуются «неинвазивные» технологии визуализации, такие как фМРТ, ПЭТ, ЭЭГ и т. Д… Возможно, вам сделали одно из таких сканирований в больнице.
Скорость передачи информации нейронами называется «скоростью нейронной передачи»; мы изучали это в эксперименте 11, когда мы измеряли скорость проводимости аксонов у дождевых червей. Однако это только одно из узких мест в скорости. Вы также должны иметь дело с синапсом (который мы изучали в эксперименте 8). Более того, скорость реакции может различаться в зависимости от того, на какой тип стимула вы реагируете и какую задачу вы выполняете.
В этом эксперименте вы и ваш друг будете проверять время реакции друг друга с помощью простой 12-дюймовой линейки. Вы будете тестировать не только визуальные стимулы, но также слуховые и тактильные стимулы.
Процедура
Этот эксперимент будет разбит на два этапа. В первом тесте будет использоваться одна линейка, а во втором тесте — две.
Эксперимент 1. На этом этапе вы и ваш партнер будете проверять время визуальной, слуховой и тактильной реакции с помощью одной линейки.
- Пусть ваш друг сядет за стол, положив доминирующую руку за край.
- Сначала проверим визуальный отклик. Держите линейку на отметке 30 см так, чтобы конец 0 см находился как раз на указательном пальце вашего друга.
- Скажите своему другу, что когда вы отпустите линейку, они должны схватить ее как можно быстрее. Не делайте никаких звуков или жестов, когда отпускаете линейку. Они должны реагировать на визуальный стимул, связанный с освобождением правителя.Запишите сантиметровую отметку.
- Повторите эксперимент еще три раза. Затем поменяйтесь местами со своим партнером и повторите это.
- Теперь вы будете записывать слуховые реакции. Пусть ваш партнер сядет за стол, как и раньше, также убедитесь, что ваш партнер надел очки.
- Опять тестируя доминирующую руку, скажите своему партнеру, что вы скажете слово «Отпустить», когда отпускаете линейку. Как только они схватят его, запишите сантиметровую отметку и повторите 3 раза. Снова поменяйтесь местами с партнером.
- Для последнего теста попросите вашего партнера снова сесть за стол в очках. На этот раз вы проверите тактильную реакцию. Скажите партнеру, что вы дотронетесь до плеча его недоминантной руки, когда отпустите линейку.
- Не подавайте партнеру никаких звуковых сигналов, которые вы отпускаете, а только простое прикосновение. Запишите измерение и, как и раньше, повторите три раза, затем поменяйте местами и повторите.
Вот таблица для первого эксперимента:
Эксперимент 2: на этом этапе вы и ваш партнер будете проверять время визуальной и слуховой реакции с помощью двух линейок.
- Для визуальной части этого эксперимента пусть ваш партнер сядет за стол, как и раньше, но обе руки должны быть за край.
- На этот раз вы будете держать обоих правителей вместо одного.
- Скажите своему партнеру, что вы отпустите только одну линейку, и он должен выбрать правильную и схватить ее как можно быстрее… Скажите им, что они не должны сжимать обе руки, только одну.
- Когда вы будете готовы начать, случайным образом выберите одну линейку.Неважно, какой именно, вы выполните этот тест еще 3 раза, но никогда не говорите своему партнеру, какую линейку вы уроните.
- Снова, как и прежде, поменяйте роли и повторите.
- Наконец, еще раз проверим слуховую реакцию. На этот раз используя обе линейки.
- Примите то же положение, что и раньше, с обеими линейками. Убедитесь, что на вашем партнере есть тени для век.
- Затем вы скажете «влево» или «вправо». Как только вы это скажете, вы опустите соответствующую левую или правую линейку.Ваш партнер должен решить, за какую линейку ухватиться, основываясь на слуховом сигнале, который вы даете: «влево» или «вправо». Как и прежде, ваш партнер должен сжимать только одну руку.
- Запишите измерение и повторите еще 3 раза, не забывая случайным образом решить, какую линейку опустить. Поменяйтесь ролями и повторите.
Вот таблица для второго эксперимента:
Математика
На приведенной выше диаграмме вы собираетесь взять все собранные вами сантиметровые измерения и преобразовать измерения из сантиметров в секунды.Это покажет вам, сколько времени в секундах требуется объекту (линейке), чтобы упасть на определенное расстояние. Приведенная ниже формула состоит из трех переменных.
Вот пример используемого уравнения:
Преобразование вручную каждого записанного числа может показаться утомительным, поэтому вместо этого вам будет предоставлена быстрая диаграмма для преобразования ваших сантиметров в секунды.Однако в таблице отсутствуют некоторые значения. Вам нужно будет их заполнить, чтобы заполнить таблицу. Используйте приведенное выше уравнение, чтобы заполнить оставшуюся часть диаграммы. Если вы сообразительны, вы также можете разработать для этого компьютерную программу.
После использования диаграммы и преобразования ваших сантиметров в секунды у вас будет время реакции линейки в секундах. Глядя на свои данные, вы можете подумать, как вы сравниваете со средним человеческим временем реакции.Вот! Среднее время реакции человека составляет 0,25 секунды на визуальный стимул, 0,17 секунды на звуковой стимул и 0,15 секунды на сенсорный стимул.
Краткая памятка для класса
Этот раздаточный материал был разработан Вирджинией Джонсон, аспирантом, которая адаптировала наш эксперимент для использования в качестве учебного пособия. Этот раздаточный материал содержит прекрасные инструкции для визуальных, тактических и звуковых экспериментов, проводимых здесь, в сжатом виде на одной странице! Спасибо, Вирджиния, что поделилась с нами!Идеи проектов для Science Fair
- Как вы думаете, почему на прикосновение и звуковые раздражители в среднем быстрее реагируют? Насколько это быстрее? Соответствуют ли ваши результаты указанным выше средним показателям?
- Ожидаете ли вы разницы в среднем времени реакции между мужчиной и женщиной? А как насчет более спортивного человека по сравнению с более сидячим человеком?
- Почему бы не проверить «тактильное» время реакции в задании выбора? Как бы вы могли переделать экспериментальную установку, чтобы проверить время тактильной реакции в задаче выбора?
- Как известно, у вас есть доминирующий vs.недоминантная рука. Всего с четырьмя попытками слишком сложно увидеть разницу. Возможно, вам стоит повторить эксперимент 10-20 раз с доминирующими правой и левой руками, чтобы увидеть, есть ли разница между доминирующими и недоминантными руками.
- Средняя скорость проводимости составляет примерно 20-80 м / с. Нейромедиатор проходит через синапсы примерно за 1 мс.
- Рассчитайте нижний предел рефлекса надколенника по сравнению с тем, что вы себе представляете для рефлекса надколенника жирафа.В качестве теста посмотрите, сможете ли вы найти разницу в скорости реакции у людей разного роста.
Измерений рефлекса
Reflex измерения
Описание
Акустический рефлекс — это сокращение стремечной мышцы, вызванное воспроизведением громкого звука. Когда в любое ухо подается громкий звук, стремительные мышцы с обеих сторон сокращаются. Сокращение стремени отклоняет переднюю стремечку от овального окна и укрепляет цепочку слуховых косточек. Это приводит к увеличению импеданса, который измеряется как небольшое снижение податливости ушным каналом.
Стременная мышца иннервируется седьмым черепным (лицевым) нервом (CNVII). Следовательно, при параличе CNVII вероятно поражение стремени.
Зачем нужно проводить измерения рефлекса?
Результаты акустического рефлекса вносят большой вклад в дифференциальную диагностику и должны быть частью каждой базовой аудиологической оценки. Они могут предоставить / подтвердить информацию о типе (кондуктивная, сенсорная, нервная) и степени потери слуха.
Акустический рефлекс, скорее всего, будет вызван при соблюдении всех следующих условий:
- Нормальная функция среднего уха
- Достаточно громкий стимул, чтобы вызвать ответ
- Нет аномальной адаптации к раздражителю
Однако примерно у 5% взрослого населения слуховые рефлексы отсутствуют.
- Диапазон интенсивности чистого тона, вызывающего акустический рефлекс, составляет от 70 до 100 дБHL (медиана = 85 дБHL)
- Ипсилатеральные АРТ у пациентов с нормальным слухом обычно на 70-80 дБ выше их пороговых значений чистого тона и примерно на 5 дБ выше их контрлатерального порога (т. Е. Если пороги чистого тона были на уровне 10 дБСП, можно было бы ожидать ипсилатеральные АРТ между 80-90 дБСП и контралатеральные АРТ. между 85-95dBHL в качестве приближения)
Противопоказатели для рефлекторной проверки
- Тиннитус
- Инфекция наружного уха
- Тяжелая вербовка
- Hyperacusis
Как настроить протокол рефлекс
- Ипси- и контралатеральный путь
Можно создать протокол для измерения либо ипсилатерального пути, либо контралатерального пути, либо обоих.Ипсилатеральный означает «та же сторона», а противопоставление означает «противоположная сторона». На рефлексы всегда указывает зонд.
Ипсилатеральный путь: Ипсилатеральный путь лучше всего можно объяснить следующим образом. Громкий звук проходит через наружное, среднее и внутреннее ухо, затем по вестибулокохлеарному нерву (CNVIII) к стволу мозга, достигая ядра улитки. Отсюда сигнал перемещается в верхний оливарный комплекс и к ядрам CNVII. Затем сигнал посылается по CNVII, вызывая сокращение стремени.
Контралатеральный путь: В контралатеральном рефлекторном пути громкий звук проходит через внешнее, среднее и внутреннее ухо, затем по CNVIII к стволу мозга, достигая кохлеарного ядра. Отсюда сигнал перемещается к другому вышестоящему оливковому комплексу и другим ядрам CNVII. Затем сигнал посылается по CNVII, вызывая сокращение стремени.
Какой пробный тон мне следует использовать?
Обычно используется пробный тон с частотой 226 Гц, если новорожденные не проходят тестирование.В этом случае используется высокочастотный пробный тон (1000 Гц).Порог акустического рефлекса (ART)
ART — это наименьшая интенсивность акустического стимула, которая вызывает результат акустического рефлекса (измеряемое изменение акустического иммитанса). Критерий изменения или отклонения 0,03 обычно принимается как минимальное изменение, необходимое для подтверждения наличия рефлекса.Тестовые частоты
Измерения ART обычно проводятся при 500, 1000, 2000, 4000 Гц.Результаты варьируются на частоте 4000 Гц, и многие молодые люди с нормальным слухом повышают уровень АРТ на этой частоте. Поэтому к результатам следует относиться с осторожностью. Некоторые врачи предпочитают использовать широкополосный шум (BBN) в качестве альтернативного стимула к 4000 Гц. Обычно шумовые стимулы вызывают рефлексы на более низком уровне, чем чистые тоны; примерно на 20 дБ ниже.Intensity
Интенсивность должна начинаться с 70-80dBHL до 105dBHL с шагом 5 дБ, пока не будет достигнут порог акустического рефлекса.В зависимости от требуемого результата тестирования (скрининговый или клинический) не рекомендуется превышать 105 дБСП, если вы не подозреваете кондуктивную потерю.
Примечание. Тестирование акустического рефлекса может вызвать необратимое повреждение слуха и шум в ушах, и, хотя стандартов для безопасных уровней презентации не существует, большая часть литературы рекомендует тестирование не выше 105–110 дБСП.Отображение положительных или отрицательных рефлексов
Рефлексы будут отображаться как положительно, так и отрицательно, в зависимости от ваших настроек.
Отрицательно отображаемый рефлекс Положительно отображаемый рефлекс

Процедура
- Предупредить клиента, что он услышит громкие звуки в любом ухе. Попросите их сесть спокойно и тихо.
- Поместите зонд иммитанса (зонд, используемый для тимпанометрии) в ухо, которое вы хотите проверить.Вставьте контралатеральный зонд в другое ухо.
- Сначала выполните тимпанометрию. Акустические рефлексы следует измерять при установленном давлении в слуховом проходе для получения максимальной податливости в присутствии зондирующего тона 226 Гц (т. Е. После тимпанометрии).
- Нажмите Старт .
Затем измерение запустит рост рефлекса и автоматически остановится при наличии рефлекса с определенными пороговыми критериями.
Если тон достаточно громкий и происходит сокращение стремени, зонд иммитанса регистрирует наличие акустического рефлекса. - Присутствие рефлекса автоматически подтверждается двойным запуском измерения с одинаковой интенсивностью для подтверждения воспроизводимости найденного порога. В качестве альтернативы, измерение можно повторить вручную на 5 дБ выше полученного ART, чтобы убедиться, что это настоящее ART.
Причины для повторных измерений рефлекса
Если во время тестирования произойдет одно из следующих событий, целесообразно провести повторный тест, чтобы подтвердить, что ваши результаты верны:
- Клиент глотает, разговаривает, смеется, кашляет и т. Д.во время теста.
- Странный результат, который не выглядит правильным или не соответствует результатам аудиограммы. В случае сомнений повторите тест, чтобы проверить повторяемость результатов.
- Суженные каналы могут привести к ложным результатам, особенно если наушники используются на контралатеральном ухе. Перепроверьте результаты, если они выглядят подозрительно или не совпадают с результатами других тестов.
Интерпретация акустических рефлексов
Рефлексы обозначаются ухом зонда, поэтому при получении рефлексов для правого уха зонд помещается в правое ухо, а противонаушники — на левое ухо.
При получении рефлексов для левого уха зонд помещают в левое ухо, а контра наушник — на правое ухо.
Какие характеристики я ищу в своих результатах?
- Наличие или отсутствие стремечного рефлекса
- порог акустического рефлекса
- Распад или адаптация акустического рефлекса (если проверено)
Ниже приводится пример того, как должен выглядеть рефлекс.Здесь значение отклонения отображается при наличии рефлекса. Однако обнаружение основано на измерении, соответствующем алгоритму. Поэтому для клинициста важно посмотреть на морфологию рефлекса в сочетании с вашей ситуацией тестирования (клиент говорит, глотать) и решить, действительно ли рефлекс является «истинным» рефлексом, а не артефактом.

Рефлекс должен показывать отклонение вниз от 0,00 мл, которое привязано ко времени предъявления стимула.Затем он будет удерживать изменение соответствия до возврата к 0,00 мл.
В примере ниже вы заметите, что рефлекс переходит в отрицательную часть графика, прежде чем перейти к положительной точке отклонения 0,05 мл. Этот тип ответа называется двухфазным и может возникать в начале или одновременно в появлении и смещении рефлекса. Аномальный паттерн двухфазного ответа как в начале, так и в конце связан с отосклерозом, особенно на его ранних стадиях.
Примеры рефлексов
Примеры нерефлексов
Почему эти образы не являются рефлексами? Во всех примерах, кроме одного (выделенного зеленым), система обнаружила, что это не рефлексы. В большинстве этих примеров клиент либо двигался, либо глотал, либо говорил во время теста, в результате которого был создан артефакт.
В первом примере, хотя это может показаться рефлексом, он не был принят как таковой, потому что не отвечал критериям отклонения, установленным прибором.
В четвертом (выделенном зеленым) примере система обнаружила это как рефлекс, но на самом деле это артефакт. По форме вы говорите, что она не соответствует образцу рефлекса. В этом случае вам следует повторить тест с этой частотой, чтобы подтвердить истинное АРТ.
Образцы акустических рефлексов
Ниже приведены девять примеров образцов рефлексов, с которыми вы можете столкнуться во время тестирования. Однако вы должны знать, что это не те результаты или закономерности, которые вы будете видеть каждый раз, когда проводите тестирование, и что клинические интерпретации в реальной жизни намного сложнее.Разные авторы по-разному публикуют шаблоны или записывают результаты, поэтому приведенные ниже таблицы являются только руководством.
Обратите внимание, что рефлексы на частоте 4000 Гц могут присутствовать или не присутствовать из-за изменчивости на этой частоте (обсуждалось ранее). Вы можете использовать BBN в качестве альтернативы тестированию на частоте 4000 Гц.
Нормальный слух и функция среднего уха
Как правило, у клиентов с нормальным слухом и нормальной функцией среднего уха ипсилатеральные и контралатеральные рефлексы будут присутствовать на всех частотах.
Пример 1: Нормальный слух / функция среднего уха
| Частота | ,5 кГц | 1 кГц | 2 кГц | 4 кГц | |
| Зонд R | Stim R (испи) | 85 | 85 | 85 | 85 |
| Stim L (против) | 90 | 90 | 90 | 90 | |
| Датчик L | Stim L (испи) | 80 | 80 | 80 | 80 |
| Stim R (против) | 85 | 85 | 85 | 85 |
Кондуктивная потеря слуха
Акустические рефлексы будут отсутствовать, если зонд помещен в ухо с заболеванием среднего уха.Это связано с тем, что при заболеваниях среднего уха датчик обычно не позволяет измерить изменение податливости при сокращении стремени. Следовательно, рефлексы будут отсутствовать даже в случае легкой кондуктивной тугоухости. При наличии тимпанограммы типа C, в зависимости от степени отрицательного давления в среднем ухе, рефлексы могут присутствовать или отсутствовать.
Если акустические рефлексы присутствуют в зондирующем ухе, маловероятно, что существует кондуктивная потеря слуха, за исключением редкого случая расхождения верхнего полукружного канала (SSCD).
Пример 2: Нормальный слух в правом ухе и умеренная кондуктивная потеря в левом ухе
| . | Частота | ,5 кГц | 1 кГц | 2 кГц | 4 кГц |
| Зонд R | Stim R (испи) | 85 | 85 | 85 | 85 |
| Stim L (против) | 100 | 100 | 100 | 105 | |
| Датчик L | Stim L (испи) | X | X | X | X |
| Stim R (против) | X | X | X | X |
В этом примере повышенные пороги левого контралатерального рефлекса (зонд справа, стимул слева) обусловлены дополнительным SPL, необходимым для преодоления легкой потери в левом ухе.Легкая патология среднего уха может влиять на сигналы, проходящие через левое ухо или измеряемые в левом ухе. Они либо отсутствуют, либо повышены.
Пример 3: Нормальный слух в правом ухе и умеренная кондуктивная потеря в левом ухе
| Частота | ,5 кГц | 1 кГц | 2 кГц | 4 кГц | |
| Зонд R | Stim R (испи) | 85 | 85 | 85 | 85 |
| Stim L (против) | X | X | X | X | |
| Датчик L | Stim L (испи) | X | X | X | X |
| Stim R (против) | X | X | X | X |
В этом примере из-за умеренной потери в левом ухе стимул (даже на максимальных уровнях) был недостаточно громким, чтобы вызвать рефлекс стремени в левой контралатеральной записи (зонд справа, стимул слева).
Кохлеарная потеря слуха
В ушах с кохлеарной тугоухостью акустический рефлекс может быть вызван на уровнях чувствительности (SL) менее 60 дБ. SL — это разница между ART и порогом слышимости. Например, если порог слышимости при 1 кГц составляет 50 дБСП, а значение ART — 90 дБСП, уровень ощущения составляет 40 дБСП.
Когда SL меньше 60 дБ, отображается положительный результат теста Metz. Это указывает на поражение улитки (нейросенсорную потерю) из-за явления набора громкости.
Пример 4: От легкой до умеренной потери улитки как в левом, так и в правом ухе
| Частота | ,5 кГц | 1 кГц | 2 кГц | 4 кГц | |
| Зонд R | Stim R (испи) | 85 | 80 | 80 | 100 |
| Stim L (против) | 85 | 85 | 90 | X | |
| Датчик L | Stim L (испи) | 85 | 90 | 85 | 100 |
| Stim R (против) | 90 | 80 | 85 | X |
В этом примере обратите внимание, что ART происходят примерно на нормальном уровне.Это связано с тем, что порог акустического рефлекса в ухе с потерей улитки может напоминать результаты для нормального уха, когда пороги воздушной проводимости ниже примерно 50 дБСП. По мере увеличения порога слышимости выше этого уровня увеличивается вероятность регистрации повышенного или отсутствующего акустического рефлекса.
Пример 5: Тяжелая или глубокая улитка в левом ухе, нормальный слух в правом ухе
| Частота | .5 кГц | 1 кГц | 2 кГц | 4 кГц | |
| Зонд R | Stim R (испи) | 85 | 85 | 85 | 95 |
| Stim L (против) | X | X | X | X | |
| Датчик L | Stim L (испи) | X | X | X | X |
| Stim R (против) | 90 | 90 | 90 | 95 |
В этом примере стимул (даже на максимальных уровнях) был недостаточно громким, чтобы вызвать рефлекс стремени из-за тяжелой / глубокой потери в левом ухе.Таким образом, всякий раз, когда в пораженное ухо подается стимул, рефлексы будут отсутствовать / повышаться как на ипсилатеральной, так и на контралатеральной записи, как показано выше.
Ретрокохлеарная потеря слуха
АРТ ушей с ретрокохлеарной патологией (CNVII) обычно выше, чем при нормальном слухе или улитковой тугоухости. Часто они отсутствуют при максимальных уровнях стимула. Имейте в виду, что результаты АРТ следует анализировать в сочетании с историей болезни пациента, аудиограммой, речевыми данными и данными тимпанометрии для дифференциальной диагностики.
Несколько замечаний:
- Уши с ретрокохлеарной патологией и нормальным слухом не имеют рефлексов в 30% случаев
- При легкой потере слуха 30 дБ вероятность отсутствия рефлексов увеличивается
- Отсутствие рефлексов на частотах 0,5, 1 и 2 кГц при нормальном / почти нормальном слухе следует рассматривать как подозрительное, если не доказано иное.
- Пораженное ухо покажет отсутствие акустических рефлексов при предъявлении к нему стимула в случае поражения CNVIII
Пример 6: Ретрокохлеарное поражение в левом ухе; нормальный слух на оба уха
| Частота | .5 кГц | 1 кГц | 2 кГц | 4 кГц | |
| Зонд R | Stim R (испи) | 80 | 80 | 80 | 90 |
| Stim L (против) | 105 | 110 | X | X | |
| Датчик L | Stim L (испи) | 110 | X | X | X |
| Stim R (против) | 85 | 80 | 85 | 95 |
В этом примере обратите внимание на повышенные / отсутствующие акустические рефлексы с направлением в левое ухо.
Пример 7: Ретрокохлеарное поражение / поражение CNVIII в левом ухе; легкая потеря слуха в левом ухе и нормальный слух в правом ухе
| Частота | ,5 кГц | 1 кГц | 2 кГц | 4 кГц | |
| Зонд R | Stim R (испи) | 80 | 80 | 85 | 85 |
| Stim L (против) | X | X | X | X | |
| Датчик L | Stim L (испи) | X | X | X | X |
| Stim R (против) | 85 | 85 | 90 | 90 |
В этом примере обратите внимание на отсутствие акустических рефлексов, когда звук подается в левое ухо.
Поражение лицевого нерва / CNVII
Акустические рефлексы отсутствуют при измерении на пораженной стороне в случае поражения лицевого нерва (например, зонд в пораженном ухе). Это связано с тем, что стременная мышца иннервируется CNVII.
Часто расстройства CNVII легко распознаются (например, паралич лицевого нерва в случае паралича Белла), и измерение акустического рефлекса используется в качестве инструмента для наблюдения за процессом выздоровления у таких пациентов.
Пример 8: Поражение лицевого нерва / CNVII в левом ухе вследствие паралича Белла; нормальный слух на оба уха
| Частота | .5 кГц | 1 кГц | 2 кГц | 4 кГц | |
| Зонд R | Stim R (испи) | 80 | 80 | 85 | 85 |
| Stim L (против) | 85 | 85 | 85 | 90 | |
| Датчик L | Stim L (испи) | X | X | X | X |
| Stim R (против) | X | X | X | X |
Обратите внимание на то, что в этом примере акустические рефлексы отсутствуют, когда зонд подсоединен к пораженному (левому) уху.Кроме того, вы узнаете, что это аналогичная картина результатов для поражения CNVIII.
Межосевое поражение ствола мозга
- Очень редко. Примерно 1 из 10 миллионов
- Акустические рефлексы нормальны на ипсилатеральном направлении и отсутствуют на контралатеральном направлении. Левый и правый проводящие пути нарушены поражением слуховых волокон.
Пример 9: Внутриаксиальное поражение ствола мозга; нормальный слух на оба уха
| Частота | .5 кГц | 1 кГц | 2 кГц | 4 кГц | |
| Зонд R | Stim R (испи) | 80 | 80 | 85 | 85 |
| Stim L (против) | X | X | X | X | |
| Датчик L | Stim L (испи) | 85 | 80 | 80 | 85 |
| Stim R (против) | X | X | X | X |
Распад рефлекса
Тестирование распада рефлекса может быть полезно для выявления / подтверждения ретрокохлеарной патологии у пациентов.Как правило, пациенты будут иметь типичные ретрокохлеарные признаки (односторонний шум в ушах, асимметричная потеря слуха, головокружение / вертиго), и у вас будет достаточно информации для направления к ЛОР-специалисту без необходимости проведения этого теста. Этот тест может быть полезен, если аудиограмма и история болезни в норме, но рефлекторные результаты показывают ретрокохлеарный паттерн.
Тест на угасание акустического рефлекса позволяет определить, сохраняется ли рефлекторное сокращение или ослабевает во время непрерывной стимуляции (обычно 10 секунд).Тестирование обычно проводится на частотах 500 Гц и 1000 Гц, но не выше этих частот, поскольку даже нормальные уши могут показывать затухание на более высоких частотах.
Тест выполняется путем подачи непрерывного стимула на 10 дБ выше ART для этой частоты в течение 10 секунд. Либо величина рефлекторной реакции останется прежней, либо уменьшится в течение 10 секунд. Что вы ищете, так это то, уменьшится ли ответ до половины своей первоначальной величины. Следовательно, если рефлекторный ответ уменьшится до 50% от своей первоначальной величины в течение 10 секунд тестирования, тест будет положительным в отношении распада рефлекса.
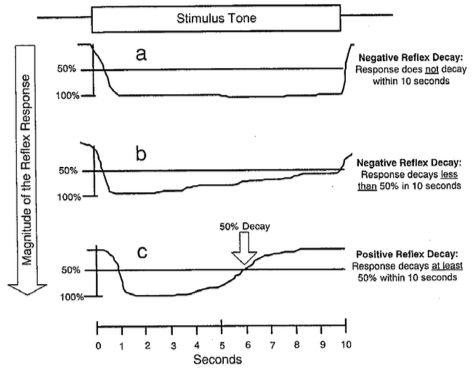
На приведенном выше рисунке (взятом из Gefland, 2001) спад акустического рефлекса считается отрицательным, если рефлекторный ответ не уменьшается (пример a) или если он уменьшается менее чем на половину от его первоначальной величины (пример b). Распад рефлекса положительный, если его величина падает на 50% или более (как в примере c).
Процедура
- Сначала выполните тимпанометрию и измерения рефлекса.
- Возьмите порог акустического рефлекса на 500 Гц или 1000 Гц в ухе, которое вы хотите проверить, и добавьте 10 дБ.
Это уровень стимула, который вы будете использовать для тестирования (например, если порог рефлекса составлял 80 дБ при 1000 Гц, вы бы проверили на 90 дБ при 1000 Гц). Интенсивность автоматически устанавливается на 10 дБ выше найденного порога рефлекса. - Убедитесь, что между зондом и ухом все еще есть хорошее уплотнение, и затем нажмите кнопку «Пуск», чтобы запустить тест.
Примечание : Если тест распада рефлекса положительный, вы должны убедиться, что это не из-за неправильной печати, которая может привести к артефакту, похожему на кривую затухания.
Интерпретация распада рефлекса
Посмотрите на записанное измерение. Обычно вы увидите результат, аналогичный приведенному ниже примеру.
Значение затухания — это процентная разница между двумя значениями отклонения рефлекса, полученными через полсекунды после начала стимула и полсекунды до его остановки.
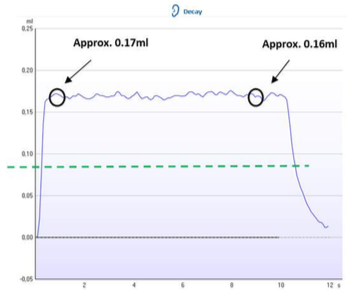 Пример 1. В этом примере тест распада рефлекса отрицательный, так как ответ не уменьшился более чем на 50% (падение ниже зеленой пунктирной линии) в течение 10-секундного интервала тестирования.Синяя линия рефлекса должна была опуститься ниже зеленой пунктирной линии, чтобы можно было измерить положительный распад рефлекса.
Пример 1. В этом примере тест распада рефлекса отрицательный, так как ответ не уменьшился более чем на 50% (падение ниже зеленой пунктирной линии) в течение 10-секундного интервала тестирования.Синяя линия рефлекса должна была опуститься ниже зеленой пунктирной линии, чтобы можно было измерить положительный распад рефлекса.
 Пример 2. В этом примере показано измерение затухания положительного рефлекса. Рефлекс снизился более чем на 50% (обозначено зеленой пунктирной линией) в течение 10-секундного интервала тестирования.
Пример 2. В этом примере показано измерение затухания положительного рефлекса. Рефлекс снизился более чем на 50% (обозначено зеленой пунктирной линией) в течение 10-секундного интервала тестирования.
Ссылки
Bess, F.H., & Humes, L.E. (2003). Аудиология: основы (3-е изд.). Балтимор: Липпинкотт Уильямс и Уилкинс.
Кэмпбелл, К.К., и Муллин Г. (2006). Импедансная аудиометрия. Проверено 2 октября 2009 г..
Эммануэль К. Д. (2009). Образцы порога акустического рефлекса (ART): руководство по интерпретации для студентов и руководителей. Проверено 2 октября 2009 г..
Фаулер, К. Г., и Шанкс, Дж. Э. (2002). Тимпанометрия. В Дж. Кац (ред.), Справочник по клинической аудиологии (5-е изд.). (стр. 175 — 204). Балтимор: Липпинкотт Уильямс и Уилкинс.
Gefland, S.A. (2001). Основы аудиологии (2-е изд.). Нью-Йорк: Thieme Medical Publishers, Inc.
Марголис, Р. Х., & Хантер, Л. Л. (2000). Измерения акустического иммитанса. В R. J. Roeser, M. Valente & H. Hosford-Dunn (Ed.), Audiology диагностика. (стр. 381 — 423). Нью-Йорк: Thieme Medical Publishers, Inc.
Сриредди, С. В., Райан, К. Э. и Нипарко, Дж. К. (2003). Обследование пациента с потерей слуха. В J. Niparko & L.
R. Lustig (Ed), Клиническая невротология: диагностика и лечение нарушений слуха, равновесия и лицевого нерва.(стр.65 — 80). Лондон: Издательство Мартина Дуница.
Зито Ф. и Роберто М. (1980). Картина акустического рефлекса изучается методом усреднения. Аудиология, 19, 395-403.
.| Срок | Определение
| ||
| Срок | Определение
| ||
Срок
| Определение
| ||
| Срок | Определение
| ||
| Срок | Определение
| ||
| Срок | Определение
| ||
Срок
| Определение
| ||
Срок
| Определение
| ||
| Срок | Определение
| ||
Срок
| Определение
| ||
| Срок | Определение
| ||
| Срок | Определение
| ||
Срок
| Определение
| ||
| Срок | Определение
| ||
| Срок | Определение
| ||
Срок
| Определение
| ||
| Срок | Определение
| ||